Мембрана нейрона в состоянии покоя. Возбуждение нейрона
1. Барьерная (защищает клетку, поддерживает ее форму)
2. Транспортная (определяет состав веществ внутри клетки)
3. Рецепторная (определяет специальную чувствительность данной клетки к определенной группе химических веществ)
4. Электрическая (обеспечивает создание разности потенциалов между внутренней и внешней поверхностью мембраны).
Отличительные свойства нервной клетки:
Возбудимость (способность генерировать потенциал действия при раздражении)
Проводимость (способность проводить и передавать возбуждение другим клеткам).
Электрические процессы в нейронах
Природа мембранного потенциала (потенциала покоя)
Потенциал покоя формируется благодаря пассивному (по градиентам) выходу ионов калия из клетки. В результате: -на наружной поверхности мембраны возникает избыток положительно заряженных ионов; внутри клетки остаются отрицательно заряженные крупные молекулы.
Механизм возбуждения нейрона:
1. Начальное изменение потенциала мембраны;
2. раскрывается часть натриевых каналов;
3. повышается проницаемость мембраны для натрия;
4. перемещение натрия в клетку по электрическому и химическому градиентам.
5. Рост числа положительны ионов внутри клетки;
6. Локальная деполяризация мембраны. (если деполяризация незначительна, то все сначала)
Если деполяризация достигает критической величины:
7. Раскрываются все натриевые каналы;
8. Происходит резкая деполяризация мембраны - потенциал действия (от -90 мВ до +30 мВ)
9. Натриевые каналы захлопываются и раскрываются калиевые каналы (через 0,5 мс).
10. Прекращается диффузия натрия, и начинается выход калия, который вытягивает электрический градиент.
11. Восстановление мембранного потенциала до исходных значений – реполяризация.
12. Прекращение выхода калия из клетки за счет изменения электрического градиента.
13. Включается натрий-калиевый насос.
14. Восстановление исходного ионного баланса (калий – внутри клетки, натрий – снаружи).
Ионный насос – мембранная транспортная система, обеспечивающая перенос ионов против электрохимического градиента, то есть с затратой энергии.
[рис. Потенциал действия]
Закон «все или ничего»
Если деполяризация мембраны достигает критической (пороговой) величины, то формируется потенциал действия. Если деполяризаця мембраны не достигает пороговой величины, то потенциал действия не формируется.
Преимущества сальтаторного проведения:
Экономичность (площадь перехвата менее 1% мембраны аксона)
Скорость (поле распространяется на большее расстояние).
Структурные элементы синапса:
1. Пресинаптическая мембрана (мембрана аксона, передающего нейрона)
2. Синаптическая щель (межклеточная жидкость)
3. Постсинаптическая мембрана (мембрана дендрита или сомы принимающего нейрона)
Механизм синаптической передачи.
1. Приход потенциала действия в синаптическое окончание аксона;
2. Раскрытие кальциевых каналов;
3. Повышение проницаемости мембраны для кальция;
4. Перемещение ионов кальция в клетку;
5. Деполяризация пресинаптической мембраны;
6. Выброс медиаторов в синаптическую щель (чем больше деполяризация – тем больше выброс).
7. Соединение медиатора со специфическими рецепторами постсинаптической мембраны;
8. Изменение потенциала постсинаптической мембраны;
9. Раскрытие ионных каналов;
10. Если увеличение проницаемости ионов натрия приводит к формированию ВПСП (возбудительный постинаптический потенциал), если калия и хлора – ТПСП.
Проведение в синапсах: одностороннее, с задержкой.
Свойства постсинаптических потенциалов:
Градуальность (амплитуда потенциалов переменная и отражает частоту потенциалов действия, поступающих на синапс)
Локальность (ВПСП и ТПСП распространяются по нейрону с затуханием).
Способность к суммации (суммируются потенциалы, близкорасположенные в пространстве и времени).
Функции нейроглии:
Защитная
Изолирующая
Обменная.
Функции глиальных клеток:
1. Астроциты: формируют каркас для нейронов; обеспечивают метаболизм; регенерация нерва.
2. Олигодендроциты: миелиновые оболочки аксонов.
Функционирование спинного мозга
Спинной мозг – это главный исполнительный отдел ЦНС. В его задачи входит передача команд на мышцы и железы, а также регуляция работы внутренних органов.
Корешки спинного мозга делятся на задние и передние.
Задние – чувствительные– афферентные. Состоят из аксонов клеток спинальных ганглиев. По ним предается информация от кожных рецепторов, проприорецепторов, висцерорецепторов.
Передние – двигательные – эфферентные. Состоят из аксоны мотонейронов. Направляются к мышцам к железам.
Каждый сегмент спинного мозга иннервирует три метамера тела.
Задние рога спинного мозга состоят из чувствительных (афферентых) нейронов, интернейронов (вставочных нейронв), а также клеток желатинозной субстанции (тормозные нейроны).
Передние рога состоят из мотонейронов.
Функциональные отделы серого вещества спинного мозга [рисунок]
По восходящим путям спинного мозга передаются:
- сигналы от рецепторов мышц и сухожилий (проприорецепторы) по пучкам Голяя и Бурдаха, по спиномозжечковым путям Говерса и Флексига.
Сигналы от болевых и тепературных рецепторов по латеральному спиноталамическому тракту.
Сигналы от тактильных рецепторов по вентральному спиноталамическому пути и частичто по пучкам Голля и Бурдаха.
Нисходящие пути спинного мозга. Включают в себя две системы: пирамидную и экстарпирамидную систему.
По пирамидной системе передается команды на выполнение целенаправленных движений, по кортикоспинальным путям.
По экстарпирамидной системе передаются команды поддержания позы и равновесия по ретикулоспинальным, руброспинальным, тектоспинальным, вестибулоспинальным и оливоспинальным путям.
Спинной мозг реализует две основных функции: рефлекторная и проводниковая.
Рефлекторная функция спинного мозга
Рефлекс – это стереотипная реакция организма на раздражение рецепторов, осуществляемая при участии нервной системы.
Дуга спинального соматического рефлекса [рисунок]
Фунзкциональной единицей спинного мозга является цепь, объединяющая чувствительный нейрон с мотонейроном.
Рефлекторная дейятельность спинного мозга обеспечиваеется :
1. передачей возбуждения с чувствительных нейронов на моторные нейроны.
2. Регуляцией передачи возбуждения в рефлекторной дуге.
Регуляция рефлексов осуществляется через организованное торможения.
1. Внутрисементарное торможение – скоординированная работа флексоров и экстензоров (сгибатели и разгибатели).
2. Межсегментарное тороможение – скоординировання работа мыщечных групп.
3. Эфферентное (центральное) торможение внутриспинальных тормозных связей – сила и скорость рефлекторной реакции.
Реализация рефлекторной функции:
1. обработка афферентных сигналов
2. обработка команд от управляющих структур
3. Формирование моторных команд
4. Формирование обратной афферентации
Рефлексы спинного мозга:
1. Мышечные (движения, поза).
2. Кожные (сосудистые, потоотделительные…)
3. Висцеральные (мышцы груди, спины…)
Спинальные рефлексы:
1. Простые (реализуются одним сегментом)
2. Сложные (реализуются несколькими сегментами)
Спинной мозг – это центр элементарных двигательных программ; центр врожденных двигательных автоматизмов;
Проводниковая функция спинного мозга
Заключается в передаче восходящих и нисходящих потоков информации. Восходящая информация – о положении конечностей, туловища, головы. Нисходящая информация содержит в себе команды на движения, поддержание позы и равновесия, регулирующие влияния (на рефлекторные дуги).
Вегетативная нервная система спинного мозга
Вегетативная нервная система – это та часть нервной системы, которая иннервирует внутренние органы, сосуды, железы и мышцы.
Состоит из двух отделов:
Симпатической системы (локализована в спинном мозге - в боговых рогах)
Парасимпатической системы (локализована в стволе головного мозга и в крестцовом отделе спинного мозга – в боковых рогах)
Центры симпатической НС
1. Грудной отдел спинного мозга (боковые рога)
2. Поясничные отдел спинного мозга (боковые рога)
Центры парасимпатической НС, находящиеся в спинном мозге, находятся в крестцовом отделе спинного мозга (боковые рога).
Нейрон ограничен липо-протеиновой(жиро-белковой) мембраной. Мембранный потенциал нейрона оказывает влияние на процессы трансмембранного обмена веществ. Мембранный потенциал покоящейся клетки называется потенциалом покоя.
Скорость диффузии веществ через мембрану зависит главным образом от размера молекул и их относительной растворимости в жирах.
В мембранах клеток существуют белки-транслоказы. Взаимодействуя со специфическим лигандом, они обеспечивают его диффузию (транспорт из области большей концентрации в область меньшей) через мембрану.
Специальные молекулярные рецепторы, находящиеся на мембране, могут соединяться с сигнальными (управляющими) веществами, вследствие чего может меняться состояние мембраны и всей клетки. Молекулярные рецепторы запускают биохимические реакции в ответ на соединение с ними лагандов (управляющих веществ).
каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её поверхностью (как внутри, так и снаружи клетки), и там они осуществляют свою ферментативную деятельность.
Как видим, основное свойство мембраны – это её проницаемость по отношению к различным веществам.
Легче и лучше всего через мембрану пассивно проникают жирорастворимые неполярные мелкие молекулы. Так, легче всего с помощью простой диффузией проходят через мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Несколько медленнее диффундируют через липидный слой малые полярные незаряженные молекулы: СО2, NH3, Н2О, этанол, мочевина. Диффузия глицерола идёт уже значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана практически непроницаема. Таким образом, свободно проникать сквозь мембрану в клетку и обратно могут только жирорастворимые вещества, способные растворяться в жировом (липидном) слое мембраны. Транспорт других веществ через мембрану требует особых механизмов. Какие же вещества необходимо протаскивать через мембрану «насильно»? Это все полярные молекулы, не растворимые в жирах: молекулы воды, ионы (электролиты), а также более крупные молекулы питательных веществ, таких как глюкоза и аминокислоты.
Для транспорта в клетку веществ, слабо способных к диффузии через липидный слой мембраны, необходимы специальные транспортные структуры.
Виды транспортных структур мембраны:
1. Ионные каналы – специальные поры (дырочки) в мембране, образованные канальными белками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь, так и наружу.
2. Транслоказы – специальные мембранные белки, облегчающие переход вещества через мембрану за счёт своего временного связывания с диффундирующим веществом. Не требуют энергии, работают в обоих направлениях в зависимости от концентрации переносимого вещества.
3. Транспортёры – белковые структуры, насильно протаскивающие определённые вещества сквозь клеточную мембрану в определённом направлении с затратами энергии. Ионные насосы – это транспортёры ионов. По способу использования энергии для своей работы транспортёры можно разделить на «симпортные» и «антипортные». Симпортные транспортёры используют совместный транспорт в одном направлении двух веществ: одно из них должно иметь большую потенциальную энергию для движения через мембрану. Например, симпорт в клетку с помощью ионов натрия глюкозы, или симпорт ионов кальция с помощью ионов натрия. Антипортные транспортёры (обменники) используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Итак, перенос веществ через клеточную мембрану происходит различными путями.
Механизмы транспорта веществ через мембрану:
1. Простая диффузия жирорастворимых (гидрофобных) веществ через жировой слой мембраны. Это пассивный процесс под действием градиента (перепада) концентрации вещества по разные стороны мембраны. (Смотрите видео: пассивный транспорт через мембрану).
2. Неуправляемая диффузия (неуправляемый пассивный перенос) водорастворимых веществ через постоянно открытые ионные каналы мембраны.
3. Управляемая диффузия (управляемый пассивный перенос) водорастворимых веществ через управляемые ионные каналы мембраны.
4. Активный транспорт водорастворимых веществ с помощью специальных белковых транспортных структур (транспортёров) за счёт использования энергии расщепления АТФ.
Одна из самых главных транспортных структур мембраны – это фермент АТФаза. АТФазы разных видов транспортируют через мембрану ионы. Они переносят их как внутрь клетки, так и, наоборот, наружу.
Название АТФаза означает, что это фермент, нацеленный на расщепление АТФ (аденозинтрифосфатаза)
6. Потенциал действия нейрона. Изменения возбудимости при возбуждении.
Потенциал действия - это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала покоя вследствие перемещения ионов в клетку и из клетки и способный распространяться без затухания. ПД обеспечивает передачу сигналов между нервными клетками, между нервными центрами и рабочими органами.
Потенциал действия - это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.
Изменение возбудимости при возбуждении. При возбуждении возбудимость изменяется по фазам.
1) фаза первичной экзальтации - возбудимость выше нормы, реакция на порошковый и подпороговый раздражитель
2) фаза абсолютной рефрактерности - ответная реакция на раздражитель отсутствует, что обусловлено инактивацией натриевых каналов
3) фаза относительной рефрактерности - возбудимость восстанавливается, и ответная реакция становится возможной только при действии раздражителя надпороговой силы, что обусловлено выходящим калиевым током
4) фаза вторичной экзальтации - ответная реакция на подпороговый раздражитель
5) фаза субнормальной возбудимости - возбудимость ниже нормы, ответ возможен на действие надпороговой силы.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na+ и одновременно проносящий внутрь два К+.
Как видно, всё очень просто, главное – открыть нужные ионные каналы. Стимул-управляемые ионные каналы открываются раздражителем (стимулом). Хемо-управляемые ионные каналы открываются медиатором (возбуждающим или тормозным). Точнее, в зависимости от того, на какие каналы (натриевые, калиевые или хлорные) будет действовать медиатор, таков будет и локальный потенциал – возбуждающий или тормозный. А медиатор как для возбуждающих локальных потенциалов, так и для тормозных, может быть одним и тем же, тут важно, какие ионные каналы будут связываться с ним своими молекулярными рецепторами – натриевые, калиевые или хлорные.
7. Нервные волокна, их классификация. Особенности проведения возбуждения по безмиелиновым и миелинизированным волокнам.

Нервные волокна – это отростки нервных клеток, окруженные оболочками из нейроглиальных клеток.
По наличию (или отсутствию) миелиновой оболочки нервные волокна делят на миелиновые и безмиелиновые.
Классификация нервных волокон по Ллойду – Ханту учитывает диаметр волокна и скорость проведения импульса, но применяется только для чувствительных волокон.
Классификация нервных волокон по Эрлангеру – Гассеру основана на исследовании составного потенциала действия смешанного периферического нерва (например, седалищного).
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Особенности распространения возбуждения по миелиновым волокнам:
1. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому и все волокно сразу не охватывается возбуждением.
2. Возбуждение распространяется с большой скоростью.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
ГЛАВНОЕ ОТЛИЧИЕ В ТОМ, ЧТО ПО МИЕЛИНОВЫМ ВОЛОКНАМ ПРОВОДИТСЯ НАМНОГО БЫСТРЕЕ.
Безмиелиновые нервные волокна - один слой швановских клеток, между ними - щелевидные пространства. Клеточная мембрана на всем протяжении контактирует с окружающей средой. При нанесении раздражения возбуждение возникает в месте действия раздражителя. Безмиелиновые нервные волокна обладают электрогенными свойствами (способностью генерировать нервные импульсы) на всем протяжении.
Миелиновые нервные волокна - покрыты слоями шванновских клеток, которые местами образуют перехваты Ранвье (участки без миелина) через каждые 1 мм. Продолжительность перехвата Ранвье 1 мкм. Миелиновая оболочка выполняет трофическую и изолирующую функции (высокое сопротивление). Участки, покрытые миелином не обладают электрогенными свойствами. Ими обладают перехваты Ранвье. Возбуждение возникает в ближайшем к месту действия раздражителя перехвата Ранвье. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом перехвате Ранвье происходит усиление нервных импульсов.
Перехваты Ранвье выполняют функцию ретрансляторов (генерируют и усиливают нервные импульсы).
8. Законы проведения возбуждения по нерву.
1)Возбуждение возможно в любом направлении
2) Проведения возбуждения возможно только по целому нерву (не поврежденному)
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва.
Закон двустороннего проведения возбуждения по нервному волокну.Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности нервного волокна.Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности).
Закон изолированного проведения возбуждения по нервному волокну.В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
9. Синапсы. Центральные и периферические синапсы.
Си́напс - место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Периферические синапсы: нервно-мышечные
нейросекреторные (аксо-вазальные)
рецепторно-нейрональные
Центральные синапсы
аксо-дендритические - с дендритами, в том числе
аксо-шипиковые - с дендритными шипиками, выростами на дендритах;
аксо-соматические - с телами нейронов;
аксо-аксональные - между аксонами;
дендро-дендритические - между дендритами;
Центральные синапсы
В нервной системе синапсы образуются между отростками разных нейронов, а также между отростками и телами клеток. Соответственно их называют:
- Аксо-аксональными,
- Аксо-дендритными,
- Аксо-соматическими,
- Дендро-соматическими,
- Дендро-дендритными.
Количество синапсов на нейроне очень большое и достигает нескольких тысяч.
 Рис.3.4. Структура аксосоматического синапса.
Рис.3.4. Структура аксосоматического синапса.
В качестве примера может быть рассмотрен аксо-соматический синапс (между аксоном одной нервной клетки и телом другой), структура которого показана на рис.3.4. Аксон, подходя к телу другого нейрона, образует расширение, называемое пресинаптическим окончанием или терминалью. Мембрана такого окончания называется пресинаптической. Под ней располагается синаптическая щель, ширина которой составляет 10-50 мкм. За синаптической щелью лежит мембрана тела нейрона, называемая в области синапса постсинаптической.
1 - аксон,
2 - синаптическая пуговка,
3 - пресинаптическая мембрана,
4 - лостсинаптическая мембрана,
5 - рецепторы постсинаптической мембраны,
6 - синаптические пузырьки с медиатором,
7 - кванты медиатора в синаптической щели,
8 - митохондрии.
СаСБ - кальцийсвязывэющий белок.
Периферические синапсы
Периферические синапсы образованы между окончаниями эфферентного нерва и мембраной эффектора. Для соматических рефлексов это синапсы между двигательным нервом и скелетной мышцей, поэтому их еще называют нервно-мышечные синапсы (рис.3.7). Благодаря форме синаптического образования они получили название концевых пластинок. Строение и основные свойства концевых пластинок в общих чертах подобны центральным синапсам. Медиатором в нервно-мышечных синапсах служит ацетилхолин, выделяемый квантами. Разрушение медиатора в синаптической шели происходит за счет фермента холинэстеразы. Рецепторы постсинаптической мембраны относятся к никотиночувствительному типу (н-холинорепепторы), конкурентно блокируются ядом кураре, что прекращает нервно-мышечную передачу. Образуемый на постсинаптической мембране медиатор-рецепторный комплекс активирует хемо-чувствительные рецепторуправляемые Na-каналы, вызывает натриевый ток внутрь клетки, деполяризацию и формирование потенциала, называемого потенциалом концевой пластинки (ПКП), являющегося аналогом ВПСП центральных синапсов.
Важнейшим отличительным свойством нервно-мышечных синапсов от центральных является высокая амплитуда ПКП , всегда превышающая критический уровень деполяризации мембраны, поэтому ПКП не требует суммации как ВПСП, а каждый ПКП ведет к генерации потенциала действия, возбуждению и сокращению мышечной клетки.
 Рис.3.7. Структура нервно-мышечного синапса.
Рис.3.7. Структура нервно-мышечного синапса.
Возможно это связано с тем, что постсинаптическая мембрана имеет многочисленные складки, существенно увеличивающие поверхность взаимодействия медиатора с рецепторами (рис.3.7).
1 - пресинаптическая концевая пластинка,
2 - синаптические пузырьки с медиатором,
3 - митохондрии,
4 - синапгическая щель с квантами медиатора,
5 - складчатая постсинаптическая мембрана,
6 - рецепторы постсинаптической мембраны,
7 - саркоплазматический ретикулум,
8 - сократительный аппарат (миофибриллы) мышечной клетки.
10. Синапсы с электрическим механизмом передачи сигнала. Свойства.
Электрический синапс, или эфапс - электрический щелевой контакт между двумя примыкающими нейронами или иными возбудимыми клетками, которая образуется в виде узкой щели, содержащей характерные только для этого типа контактов элементы
Что такое электрический синапс? Электрические синапсы – это межклеточные образования, с помощью которых обеспечивается передача импульса возбуждения. Данный процесс происходит за счет появления электрического тока между двумя отделами под названием пресинаптический и постсинаптический.
Важная работа синапсов электрического типа Свойства электрических синапсов заключаются в следующем:
Быстрое действие (гораздо превосходит активность в синапсах химического типа); ----слабые следовые эффекты (суммации от последовательных импульсов практически нет);
Надежная передача возбуждения;
Высокая пластичность;
Передача в одну и в обе стороны.
Особенности структуры.

11. Синапсы с химическим механизмом передачи сигнала. Свойства.
Химический синапс - особый тип межклеточного контакта между нейроном и клеткой-мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество.
Синаптические контакты могут быть между аксоном и дендритом (аксодендритические), аксоном и сомой клетки (аксосоматические), аксонами (аксоаксональные), дендритами (дендродендритические), дендритами и сомой клетки.
В покое медиатор попадает в синаптическую щель постоянно, но в малом количестве. Под влиянием пришедшего возбуждения количество медиатора резко возрастает. Затем медиатор перемещается к постсинаптической мембране, действует на специфические для него рецепторы и образует на мембране комплекс медиатор-рецептор. Данный комплекс изменяет проницаемость мембраны для ионов К+ и Na+, в результате чего изменяется ее потенциал покоя.

Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов – потенциалов действия (ПД). Этот процесс можно сравнить с передачей информации с помощью включения и выключения фонарика (ПД = «вспышка света»).
Но для того, чтобы фонарик работал, нужна батарейка – источник электрической энергии. В случае нейрона таким источником является постоянный внутриклеточный заряд – потенциал покоя (ПП).
Нервные клетки ограничены липопротеиновой мембраной, являющейся электрическим изолятором. Между содержимым клетки и внеклеточной жидкостью существует разность потенциалов, так называемый мембранный потенциал.
В клетке как в единой системе все части - цитоплазма, ядро, органоиды - должны удерживаться вместе. Для этого в процессе эволюции развилась клеточная мембрана, которая, окружая каждую клетку, отделяет ее от внешней среды. Наружная мембрана защищает внутреннее содержимое клетки - цитоплазму и ядро - от повреждений, поддерживает постоянную форму клетки, обеспечивает связь клеток между собой, избирательно пропускает внутрь клетки необходимые вещества и выводит из клетки продукты обмена. Строение мембраны у всех клеток одинаково. Ее толщина составляет приблизительно 8 нм (1 нм =10Ý(-9)м).
Основу мембраны составляет двойной слой молекул липидов, в котором расположены многочисленные молекулы белков. Одни белки находятся на поверхности липидного слоя, другие пронизывают оба слоя липидов насквозь. Специальные белки образуют тончайшие каналы, по которым внутрь клетки или из нее могут проходить ионы калия, натрия, кальция и некоторые другие ионы, имеющие небольшой диаметр. Однако более крупные частицы через мембранные каналы пройти не могут.
В состоянии покоя наружная поверхность клетки всегда электроположительна по отношению к внутренней, т.е. поляризована. Эта разность потенциалов, называется потенциалом покоя, или мембранным потенциалом (МП). В образовании потенциала принимают участие 4 вида ионов: катионы натрия (положительный заряд), катионы калия (положительный заряд), анионы хлора (отрицательный заряд), анионы органических соединений (отрицательный заряд). Во внеклеточной жидкости высока концентрация ионов натрия и хлора, во внутриклеточной жидкости – ионов калия и органических соединений. В состоянии относительного физиологического покоя клеточная мембрана хорошо проницаема для катионов калия, чуть хуже для анионов хлора, практически непроницаема для катионов натрия и совершенно непроницаема для анионов органических соединений.
В покое ионы калия без затрат энергии выходят в область меньшей концентрации (на наружную поверхность клеточной мембраны), неся с собой положительный заряд. Ионы хлора проникают внутрь клетки, неся отрицательный заряд. Ионы натрия продолжают оставаться на наружной поверхности мембраны, еще больше усиливая положительный заряд.
Электрические процессы в клетках обусловлены неравномерным распределением ионов по обе стороны клеточной мембраны.
Соединение в живых существах органической материи и металлов, использование живой клетки в качестве проводника электрического тока не могут не удивлять. Чем больше подробностей о своем теле я узнаю, тем чаще возникает удивление от простых его действий. Сгибание руки может восприниматься не как привычный, почти не замечаемый процесс, а как результат слаженной работы мышц и сухожилий, получивших команду от мозга, переданную по нервным волокнам с помощью электричества. Разглядывая полосу глубоко переливающейся синевы, возникающей на куске полированного лабрадорита, который я верчу в руках, я воспринимаю излучаемые им электрические волны рецепторами глаз. В рецепторах эта информация кодируется в виде череды электрических импульсов. Она передается в головной мозг, обрабатывается им, а результат всего этого — восприятие таинственно-красивого камня, возникающие при этом эмоции. Матрица, короче:)
Вместе с восхищением возникает и желание узнать — как это все происходит, желание вникать все глубже в подробности каждого процесса.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.

Достаточно взглянуть на картинку слева, чтоб увидеть основное отличие потенциала покоя от потенциала действия: когда мембрана находится в состоянии покоя, на всей ее протяженности снаружи существует положительный электрический заряд, а внутри — отрицательный.

А потенциал действия, проходящий по нейрону от тела клетки до конца аксона, представляет собой изменение внешнего и внутреннего зарядов клеточной мембраны на небольшом участке нейрона: на очень краткий промежуток времени отрицательный и положительный заряды по разным сторонам мембраны меняются местами.
Возникают вопросы: что запускает потенциал действия? Какова его физическая природа, что именно происходит в клетке при его существовании? Каким образом он распространяется? Почему он идет только в одном направлении? Как клетка восстанавливает исходный потенциал покоя после прохождения по ней волны возбуждения?
Итак, первый вопрос: как возникает потенциал действия? Так как передача сигналов от нейронов-источников к нейрону-получателю, или постсинаптическому нейрону (postsynaptic neuron ), происходит в синапсах, запускающий потенциал действия сигнал стоит искать именно там. Через синапс в нейрон приходят сигнальные вещества, но их поступление не обязательно означает запуск возбуждения нейрона. Если бы нейрон реагировал на очень малые количества сигнальных веществ, это было бы так же неудобно, как иметь супер-чувствительный выключатель, срабатывающий, если на него случайно чихнуть.

Необходимо достаточно сильное воздействие определенных сигнальных веществ, чтобы запустить потенциал действия. В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы — временные изменения зарядов мембраны, вызываемые пришедшими через синапс сигнальными веществами .
Постсинаптические потенциалы сильно отличаются от потенциала действия по своей природе. Если потенциал действия имеет одну строго определенную величину, то величина постсинаптических потенциалов зависит только от вида и количества вызвавших их химических веществ, переданных через синапсы. В то время как потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно отличаются по величине и продолжительности. В одном синапсе нейрона приходящий нервный импульс может вызвать изменение мембранного потенциала на 0,1 мВ, а в другом — на 20 мВ. Кроме того, существуют как вызывающие потенциал действия, так и тормозящие его сигналы.
Тело клетки служит своеобразным калькулятором, простейшим аналоговым компьютером, суммирующим все поступающие в него сигналы в виде возникающих постсинаптических потенциалов. Аналоговые компьютеры устроены очень просто. Например можно построить водяной аналоговый компьютер. Если из одной трубы в банку вливается три килограмма воды, а из другой в ту же банку — пять, то взвесив банку и выяснив, что весы показывают вес, равный восьми килограммам, мы делаем вывод, что 3+5=8. На заре развития компьютерной техники использовались и водяные, и прочие аналоговые вычислительные машины, но наиболее продвинутой была, конечно, машина, которая в точности как и нейроны суммирует электрические токи. Операторы таких ЭВМ занимались вычислениями, перетыкая штекеры из одного гнезда в другое, сливая вместе разные токи.
Так вот в нейроне происходит такое же суммирование электрических токов. В процессе этого суммирования может быть перейден порог возбуждения (excitation threshold ): будет достигнуто определенное значение мембранного потенциала, которое запускает потенциал действия. Обрати внимание на то, что для достижения порога возбуждения мембранной потенциал должен увеличиваться. Если мембранный потенциал покоя нейрона равен -70 мВ, то его порог возбуждения может быть равен -55 мВ. Снижение мембранного потенциала например до -90 мВ приводит к тому, что для преодоления порога возбуждения требуется более сильное воздействие, ведь он остается прежним: -55мВ.

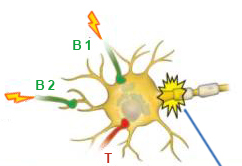
В постсинаптическом нейроне, получающем сигналы от двух возбуждающих и одного тормозящего нейронов, могут существовать различные комбинации сигналов, и далеко не каждая из них приводит к возникновению потенциала действия.
В первой схеме картинки вверху изображена ситуация, когда только от одного возбуждающего нейрона приходит слабый сигнал, которого недостаточно для преодоления порога возбуждения. Этот сигнал повторяется, но к моменту прихода второго, такого же слабого сигнала от того же аксона возбуждение мембраны уже достигает потенциала покоя, и потенциал действия не возникает.
На второй схеме изображен тот же самый слабый сигнал, но приходящий с большей частотой. Именно из-за увеличения частоты сигнала мембрана не успевает вернуться в состояние покоя в промежутке между двумя сигналами. В этом случае сумма остаточного заряда мембраны после первого сигнала и поступающего следом второго сигнала позволяет преодолеть порог возбуждения нейрона. В этом случае у основания аксона, которое называется аксонным холмиком (axon hillock [‘aksɒn ‘hɪlək] ), возникает потенциал действия.
На третьей схеме от обоих возбуждающих нейронов приходят слабые сигналы, и их суммы достаточно для запуска потенциала действия.
Четвертая схема показывает воздействие на постсинаптический нейрон возбуждающего и тормозящего сигнала. При поступлении только возбуждающего сигнала мембранный потенциал незначительно увеличивается, а при поступлении только тормозного сигнала — снижается. Если эти два сигнала приходят одновременно, то они всего лишь уничтожают друг друга, оставляя мембранный потенциал в состоянии, близком к потенциалу покоя. Это напоминает надавливание на одну и ту же дверь с разных сторон: по отдельности каждое воздействие могло бы приоткрыть дверь или наоборот, прикрыть ее, но существующие одновременно, они приводят лишь к небольшому сдвигу ее по направлению действия большей по модулю силы.
Природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект. Нейрон «складывает» все поступившие в него возбуждающие постсинаптические потенциалы, «вычитает» поступившие тормозные постсинаптические потенциалы, и как только будет перейден порог возбуждения, возникает потенциал действия.
Чтобы разобраться, что происходит в клетке во время запуска и прохождения потенциала действия , необходимо перечислить каждого участвующего игрока и его действие, как было сделано при описании возникновения мембранного потенциала покоя в . Итак, действующие лица при создании потенциала действия :

1 . , которые были закрыты при существовании мембранного потенциала покоя, открываются, как только потенциал достигает порога возбуждения. Так как внутри клетки на мембране существует отрицательный заряд, то в этот момент через каналы туда врываются притягиваемые им
2 . положительно заряженные ионы натрия (Na +). Именно они создавали положительный заряд на внешней поверхности мембраны нейрона, и именно в этот момент они переносят его внутрь, оставляя на внешней мембране отрицательный заряд, который создают находящиеся вне клетки отрицательно заряженные ионы хлора.
3 . таким образом ионы хлора (Cl —) создают отрицательный заряд на внешней поверхности мембраны.

Перемещение ионов натрия внутрь клетки приводит к резкому изменению зарядов внутри и снаружи нейрона. Место существующего во время покоя нейрона отрицательного внутреннего заряда мембраны занимает положительный заряд ионов натрия. А снаружи клеточной мембраны после ухода оттуда Na + наконец-то может проявить себя Cl — , чей отрицательный заряд больше не компенсируется положительно заряженными ионами. Таким образом потенциал клеточной мембраны внезапно возрастает с -70 до +40 мВ. Именно это повышение, называемое деполяризацией (depolarization ), изображено резко идущей вверх частью пика на этой картинке. Такое повышение мембранного потенциала является сигналом закрыть потенциал-зависимые натриевые каналы, а также открывает потенциал-зависимые калиевые каналы, о которых раньше мы еще не говорили.
4 . потенциал-зависимые калиевые каналы. Они существуют в нейроне в дополнение к проточным калиевым каналам, которые, как ты наверное помнишь из прошлой главы про потенциал, не имеют ворот и обеспечивают постоянный отток калия из клетки, играя существенную роль в формировании потенциала покоя. Так вот оказывается, что есть две разновидности калиевых каналов, и сейчас мы рассмотрим работу потенциал-зависимого калиевого канала. Открываясь после того, как потенциал действия достиг своего пика, потенциал-зависимые калиевые каналы резко увеличивают отток ионов калия из клетки. Существующий в этот момент внутри нейрона положительный заряд, образовавшийся за счет притока Na + , активно выпихивает положительно заряженные ионы калия (К +) из клетки через оба вида каналов.
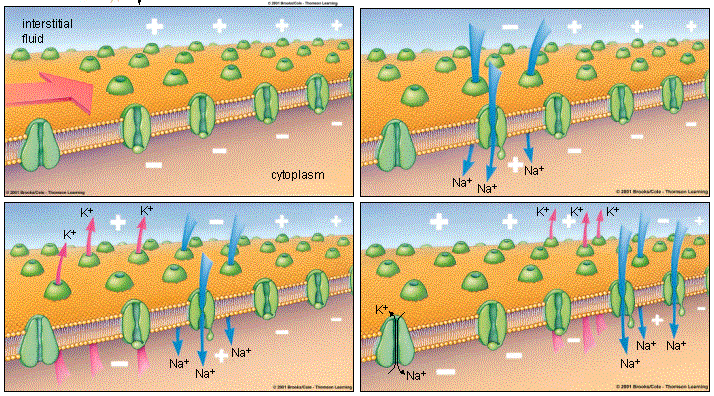
Итак, еще раз проследим цепочку событий до этого момента: потенциал достигает порога возбуждения, и этот электрический сигнал приводит к тому, что открылись потенциал-зависимые натриевые каналы, ионы Na + хлынули внутрь, притягиваемые отрицательно заряженными протеинами, произошла деполяризация, то есть разница зарядов снаружи и внутри резко снизилась, и даже более того — отсутствие положительно заряженных ионов натрия привело к возникновению отрицательного заряда там, где только что был положительный — произошла смена полюсов. Действие электрического импульса закончилось, натриевые каналы вновь закрылись, и столпившиеся внутри ионы Na + вытолкнули одноименно заряженные ионы К + через открывшиеся потенциал-зависимые калиевые каналы и через проточные калиевые каналы.
Заряд внутренней поверхности клеточной мембраны начинает стремительно снижаться, на схеме этому соответствует идущая вниз часть пика. Такое снижение заряда называется реполяризацией (repolarization ), то есть разница зарядов вне и внутри нейрона вновь восстановилась, как и было изначально в состоянии покоя. Но восстановилась именно разница зарядов, а не ионный состав, ее создающий. Несмотря на то, что в конце реполяризации снаружи мембраны образуется положительный заряд, равный положительному заряду мембраны в состоянии покоя, до достижения мембраной состояния покоя еще далеко. Ведь этот положительный заряд принесли туда вышедшие из клетки ионы К + , а при потенциале покоя его поддерживают ионы Na + , которые в данный момент заперты внутри клетки, в том самом месте, где ранее в состоянии покоя находились ионы калия.
В конце реполяризации может возникнуть снижение мембранного потенциала даже ниже его значения для покоящегося нейрона, или гиперполяризация (hyperpolarization [ˌhaɪpəpələraɪ’zeɪʃn] ). Через короткий промежуток времени она компенсируется дополнительным выходом ионов калия из клетки, и наступает прежнее, стабильное состояние покоя.

Когда мембранный потенциал становится равен потенциалу покоя (-70 мВ), потенциал-зависимые калиевые каналы закрываются. Через проточные калиевые каналы К + уже не вытекает так стремительно, ведь его концентрация внутри клетки упала, а снаружи находится много ионов калия. В то же время внутри находится много ионов натрия, которые должны быть выведены из клетки для того, чтоб через нее мог снова пройти потенциал действия.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na + и одновременно проносящий внутрь два К + . О его работе было рассказано в предыдущей главе про потенциал, и он схематично показан в четвертой части этой картинки. Совместные действия натрий-калиевого насоса и проточных калиевых каналов приводят к тому, что с внешней стороны мембраны опять возникает большая концентрация Na + , а концентрация К + внутри нейрона выше наружной.
Если бы работали только проточные калиевые каналы, то ионы калия только выходили бы из клетки, и было бы невозможным поддерживать внутри большую концентрацию К + , чем снаружи. Если бы действовали только потенциал-зависимые натриевые каналы, то ионы натрия вообще не могли бы покинуть клетку после того, как вошли туда, создавая потенциал действия. Но натрий-калиевый насос (ну то есть натрий-калиевый канал) решает обе эти проблемы: он выводит наружу Na + , восстанавливая его высокую концентрацию снаружи. И загоняет внутрь прошедшие через проточные каналы К + , не давая им возможности распределиться с обеих сторон мембраны равномерно, лишившись таким образом парциального давления, необходимого для формирования мембранного потенциала покоя.
Через тысячную долю секунды после того, как реполяризация нейронной мембраны достигла минимума, активная работа мембранных каналов восстанавливает потенциал покоя. Нейрон готов к проведению следующего потенциала действия.
5 . положительно заряженные ионы кальция (Ca 2+) играют важную роль в формировании потенциала действия лишь в некоторых нервных клетках, поэтому в данной главе, содержащей общую для различных нейронов информацию, она не будет описана.

На этом общее описание перемещения ионов через клеточную мембрану, превращающее потенциал покоя нейрона в потенциал действия, закончено. Можно сделать перерыв и переварить его вместе с чем-нибудь вкусным, заодно подпитав мозг, компенсировав те калории, которые он затратил на работу по обработке всей этой информации.
Распространение потенциала действия по всей длине аксона обеспечивается тем фактом, что возникший при его существовании в одном участке мембраны нейрона электрический ток возбуждает соседние участки, открывая натриевые потенциал-зависимые каналы и запуская уже там потенциал действия. Точнее, электрический ток запускает потенциал действия только с одной стороны от места существования предыдущего потенциала. Потому что с другой стороны натриевые каналы инактивированы после недавнего открытия, а без их участия возникновение потенциала действия невозможно. Если бы не было этого временного промежутка нефункциональности между двумя открытиями натриевых каналов, потенциал действия не мог бы распространяться только в одном направлении. Происходящая в результате цепная реакция открытия натриевых каналов обеспечивает передачу нервного импульса от места его возникновения до окончаний аксона, подходящих к другим нейронам (или к мышечной клетке).

Схематично эту цепную реакцию можно изобразить так:
Момент времени 1 : Электрический ток, возникший на в результате открытия потенциал-зависимых натриевых каналов на участке 1 , возбуждает соседние участки мембраны.
Момент времени 2 : В результате этого возбуждения на участке 2 открываются потенциал-зависимые натриевые каналы и возникает потенциал действия, который, в свою очередь, возбуждает соседние участки мембраны. На участке 1 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал после прохождения потенциала действия.
Момент времени 3 : Электрический ток, возникший в результате открытия потенциал-зависимых натриевых каналов на участке 2 , вызвал открытие таких же каналов на участке 3 , и потенциал действия перешел туда. На участке 1 потенциал-зависимые натриевые каналы не смогли открыться, несмотря на электрическое возбуждение от участка 2 , так как они были временно инактивированы после создания потенциала действия. На участке 2 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал покоя после прохождения потенциала действия.
Мы уже знаем, что отростки нейронов могут быть покрыты обернутыми вокруг них клетками нейроглии, или миелиновой оболочкой. Именно она делает аксон похожим на вытянутую цепочку сосисок, про ее образование и функции говорилось в . Передача потенциала возбуждения по «голым», не покрытым миелиновыми клетками, нервным волокнам отличается от его прохождения по миелинизированному аксону. При сравнении заметны существенные преимущества, предоставляемые обернутыми вокруг аксона клетками нейроглии:

1 . Экономичность . Мембрана аксона под миелином не имеет каналов для пропускания ионов, создающих потенциал действия. Каналы существуют только в узких промежутках между глиальными клетками, которые называют перехваты Ранвье (Ranvier’s constriction ), и только на этих узких участках аксона возникает потенциал действия. Концентрация ионных каналов в этих перехватах в 100 раз выше, чем в мембранах безмиелиновых волокон, тем не менее для того, чтобы обеспечить их работу, требуется значительно меньше энергии по сравнению с тем, сколько энергии потребовалось бы для обеспечения ионных каналов, расположенных по всей поверхности «голого» аксона. Кроме того, такая экономичность позволяет быстрее «перезаряжать» аксон для проведения следующего потенциала действия, поэтому миелинизированные волокна способны передавать сигналы более высокой частоты.
2
. Скорость
. По «голому» аксону сигнал передается в каждой его точке. Ионные каналы открываются в каждой точке его поверхности, один за другим, что напоминает тщательное, скрупулезное вчитывание в текст.
В миелинезированном аксоне потенциал действия, возникший в одном перехвате Ранвье, своим электрическим полем дотягивается до соседних перехватов, что приводит к возникновению в них новых потенциалов действия. То есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает следующий после поврежденного перехват, а в случае необходимости даже третий, четвертый и пятый по счету. Распространение электрического поля на такую длину возможно из-за электроизоляции, создаваемой миелиновыми муфтами: она уменьшает рассеивание электрического поля, которое неизбежно для безмиелиновых волокон.
Таким образом, скорость распространения потенциала действия по миелинизированным волокнам намного выше по сравнению с немиелинизированными. Ведь за один и тот же отрезок времени потенциал действия на «голом» аксоне возбудит лишь находящийся непосредственно рядом участок, а потенциал действия на миелинизированном волокне перепрыгнет на расстояние, равное расстоянию между одним или несколькими перехватами Ранвье:


Диапазон скоростей передачи потенциала действия в различных миелинизированных волокнах очень широк: от нескольких метров в секунду до «мирового рекорда», установленного аксоном креветки: он проводит возбуждение со скоростью, превышающей 200 м/с.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения потенциала действия за счёт миелиновой оболочки, достаточно сравнить скорость его распространения по немиелинизированным и миелинизированным волокнам нейронов человека. Если взять скорость проведения импульса в одинаково тонких волокнах, то в безмиелиновом волокне она составит примерно 1 м/с, а в миелинизированном 15-20 м/с. В толстых аксонах человека, покрытых миелиновой оболочкой, скорость проведения может достигать 120 м/с.
Итак, подведем итог : передача электрического импульса через тело и отростки нейрона осуществляется с помощью прохождения ионов через клеточную мембрану.

1 . сначала на клеточной мембране создается потенциал покоя . Его образуют положительно заряженные ионы натрия (Na + ) снаружи клетки и отрицательно заряженные протеины внутри клетки. Создающие заряды частицы расположены очень близко к клеточной мембране, ведь они имеют разные знаки и притягиваются друг к другу. Поэтому потенциал покоя существует только возле мембраны, в целом среда внутри нейрона и вне его электрически нейтральна. Кроме Na + и отрицательно заряженных протеинов, в создании и поддержании электрического заряда участвуют положительно заряженные ионы (К + ) и отрицательно заряженные ионы хлора (Cl — ) , тоже находящиеся вблизи мембраны. Потенциал покоя равен примерно -70 мВ (в дальнейшем при указывании численного значения мембранных потенциалов слово «примерно» будет опускаться).
2 . когда нейрон через синапсы получает сигналы от других нейронов, под их влиянием на мембране тела нейрона начинают возникать постсинаптические потенциалы , то есть изменения величины электрических зарядов на внутренней и внешней поверхностях мембраны. Под влиянием этих сигналов внутренний отрицательный заряд и внешний положительный могут увеличиваться или уменьшаться на разные величины. И нейрон выступает в роли калькулятора, суммирующего эти воздействия.
3 . итогом суммирования постсинаптических потенциалов может являться продолжение нахождения нейрона в состоянии покоя. Но если сумма дойдет до значения, называемого порогом возбуждения , то в аксонном холмике (утолщении тела нейрона у основания аксона) возникнет потенциал действия .
4 . в создании потенциала действия главную роль играет клеточная мембрана . Она регулирует прохождение через нее ионов (всех перечисленных, кроме отрицательно заряженных протеинов, которые не могут покидать пределы клетки), открывая и закрывая специализированные каналы, способные проводить только определенные ионы.
5 . когда мембрана нейрона переходит порог возбуждения, в ней открываются потенциал-зависимые натриевые каналы . Исчезает то препятствие, которое отделяло положительно заряженные Na + от притягивающих их отрицательно заряженных протеинов, и Na + устремляются внутрь клетки . Таким образом наружный положительный заряд переносится внутрь клетки, а снаружи остается отрицательный заряд, создаваемый ионами Cl — . Происходит изменение мембранного потенциала от потенциала покоя, равного -70 мВ, до +40 мВ, и это называют потенциалом действия . Важно понимать, что это изменение потенциала происходит только на том участке мембраны, где открылись потенциал-зависимые натриевые каналы.
6 . В момент достижения мембраной потенциала в 40 мВ потенциал-зависимые натриевые каналы закрываются, и на этом же участке открываются потенциал-зависимые калиевые каналы . Снаружи в этот момент существует отрицательный заряд, а внутри — положительный, создаваемый вошедшими туда ионами Na + и уже находящимися там в большом количестве ионами К + . Ионы калия выпихиваются в окружающую нейрон среду через открывшиеся каналы, что резко снижает внутренний заряд мембраны и увеличивает внешний. По обеим сторонам мембраны создаются заряды той же величины, которые обычно поддерживают потенциал покоя. Но это состояние клетки отличается от потенциала покоя тем, что ионы калия и натрия находятся не на своих местах: Na + находится внутри, а К + снаружи. Опять же все описанное касается только одного места на мембране, в котором только что перестал существовать потенциал действия.
7 . Чтобы переместить положительно заряженные ионы Na + и К + по нужные для поддержания потенциала покоя стороны мембраны, используется натрий-калиевый насос . И это действие тоже касается ограниченного участка мембраны: того, где только что закрылись потенциал-зависимые калиевые каналы.
Один цикл работы натрий-калиевого насоса заключается в том, что он захватывает изнутри клетки три иона Na + , перемещает их наружу, где захватывает два иона К + и перемещает их внутрь . То есть канал обменивает три иона Na + на два иона К + , причем Na + могут перемещаться только наружу, а К + только внутрь клетки. Таким образом восстанавливается исходная концентрация натрия на наружной стороне мембраны и калия на внутренней. Натрий-калиевый насос выводит из клетки три положительно заряженных иона, а вводит в нее только два. Следовательно, при каждом его цикле внутренний положительный заряд мембраны уменьшается, а внешний увеличивается. К тому же и вытекающие ионы калия уносят изнутри наружу свои положительные заряды. Возникает вопрос: если потенциал мембраны уже и так был равен потенциалу покоя после входа ионов калия внутрь клетки, зачем его снижать еще сильнее? Ответ заключается в том, что кроме упомянутых здесь ионов, существуют и другие, чье участие в формировании потенциала покоя менее существенно, и именно для компенсации их влияния необходимо такое выравнивание мембранного потенциала. Итогом этого процесса является достижение мембранного потенциала покоя: заряд внутренней стороны мембраны отрицателен, внешней положителен, разность потенциалов равна -70 мВ.

8 . Итак, в том месте клеточной мембраны, где всего три шага назад существовал потенциал действия, опять царит потенциал покоя. Но потенциал действия не прошел бесследно: его существование (описанное в пункте 5) создало электрическое поле, возбудившее соседние с ним части мембраны . С одной стороны (по направлению к концу аксона) это возбуждение вызвало открытие потенциал-зависимых натриевых каналов, запустив уже на новом участке потенциал действия. Который в свою очередь порождает электрическое поле, запускающее следующий потенциал действия, и этот цикл повторяется по всей длине аксона, от его основания до концов, просоединенных к следующим в нервной цепи клеткам.
С другой стороны от потенциала действия, ближе к телу нейрона, несмотря на воздействие электрического поля потенциал действия не возникает. Потому что потенциал-зависимые натриевые каналы на этом участке совсем недавно открывались, и на некоторое время они инактивированы. Именно эта пауза в их работе обеспечивает распространение потенциала действия только в одном направлении.
9 . Любой сигнал, будь то передача информации от сенсорного нейрона в мозг, или команда мозга, идущая к мотонейрону, передается одинаковыми по величине потенциалами действия . Результат потенциала действия (появление восприятия запаха, обусловленное работой сенсорных нейронов, или перемещение тела поближе к его источнику в результате слаженной работы мотонейронов) зависит исключительно от того, в какую нервную цепь включен нейрон, этот потенциал передающий. Для передачи интенсивности сигнала (силы запаха или величины сокращения мышцы) используется частота, с которой потенциалы действия передаются по нейрону. Слабый сигнал передается редкими, а сильный — частыми потенциалами действия .
10 . миелиновая оболочка вокруг отростков нейрона повышает скорость распространения потенциала действия за счет того, что сигнал передается на большое расстояние, равное ширине одной обернутой вокруг отростка клетки нейроглии, а не на непосредственно прилегающий к возбужденному участок, как это происходит у безмиелиновых волокон.
1.1. Строение нейрона
По современным представлениям нервная клетка (нейрон) представляет собой основной функциональный элемент нервной системы, перерабатывающий поступающую к нему информацию и передающий результат переработки другим нейронам.
1.1.1. Сома, дендриты, аксон
Нейрон состоит из тела клетки - сомы, множества ветвящихся отростков - дендритов, по которым поступают сигналы, и обычно одного выходного волокна - аксона. Внутриклеточная среда отграничена от внеклеточной тонкой оболочкой - мембраной.
На мембране сомы и дендритов расположены синапсы - окончания аксонов других нейронов. Сома нейрона состоит из ядра, содержащего генетический аппарат, и цитоплазмы, в которой осуществляется синтез белков. В теле нейрона, в дендритах и аксонах имеются мнкротрубочки диаметром около 200 А, которые предположительно служат для транспорта синтезируемых в теле нейрона белков.
Переход сомы нейрона в аксон называется аксонным холмиком и характеризуется высокой возбудимостью. Аксоны некоторых нейронов нервной системы покрыты оболочками, состоящими из шванновских клеток или клеток глии. Промежутки между ними - перехваты Ранвье (рис. 1.1).
Разные типы нервных клеток характеризуются разной геометрической структурой дендритов. Встречаются дендриты довольно длинные, простирающиеся от сомы на расстояние в 30-40 диаметров сомы, маловетвящиеся. Другие дендриты - на расстояние порядка диаметра сомы и имеют большое количество узлов ветвления.
1.1.2. Структура мембраны нейрона
Мембрана толщиной 70-80 А, отделяющая цитоплазму нейрона от внешней среды, состоит из слоя фосфолипидных молекул, имеющего с внешней и с внутренней сторон слои белковых молекул. Центральный фосфолипидный слой обладает симметричной структурой, в которой жирные кислоты липидов обращены внутрь мембраны.
Поперечник мембраны пронизывают каналы, по которым могут перемещаться определенные ионы. В мембрану встроены рецептивные белки, управляющие состоянием ионных каналов (рис. 1.2). В модели мембраны совокупность каналов представлена сопротивлением, а совокупность диэлектрических участков - параллельно включенной емкостью.
Мембрана, окружающая нервную клетку (сому, дендриты и аксон), бывает двух типов: пассивная и активная.
Пассивная мембрана способна проводить электрическое возбуждение лишь электротонически (с затуханием), как кабель. Активная мембрана способна формировать и проводить незатухающий с расстоянием импульс. Эти два типа мембран могут перемежаться.
Установлено, что в активной мембране существуют особые участки - каналы, которые могут быть в открытом или закрытом состоянии. Обычно каналы бывают двух типов: проводящие ионы натрия, проводящие ионы калия.
Каналы каждого из этих типов проводят только ионы одного вида (либо только калия, либо только натрия). Управление (открытие, закрытие) каналов осуществляют многовалентные катионы, в основном кальция. В невозбужденной мембране катионы кальция находятся в связанном состоянии с молекулами, образующими каналы, закрывая каналы. Смещение полярных участков некоторых молекул мембраны открывает каналы для ионов натрия и калия, что приводит к возбуждению мембраны и генерации распространяющегося потенциала действия.
1.1.3. Синапсы, типы синапсов
Синапсы - участки взаимодействия аксонных термина- леи с мембраной нейрона - бывают двух типов: возбуждающие и тормозные. Возбуждающие увеличивают деполяризацию мембраны, тормозные - гиперполяризацию мембраны.
По принципу своего влияния на мембрану синапсы подразделяются на химические и электрические. Кроме того, различают аксо-аксональные, аксосоматические, аксодендритические, дендродендритические и соматосоматические синапсы (рис. 1.3).
Хотя плотность распределения синапсов на соме и дендритах в среднем одинакова, однако более высокая плотность распределения синапсов имеет место в узлах ветвления и в области локальных расширений дендритов.
Поэтому узлы ветвления дендритов имеют особое значение при функционировании нейрона.
На соме нервных клеток в большинстве случаев синапсы мелкие, их линейные размеры не превышают 0,2- 0,4 мкм (для нервной системы млекопитающих), в то время как на дендритах значительная часть синапсов имеет протяженность около 1 мкм.
На дендритах бывают синаптические контакты специального вида, так называемые «шипиковые» синапсы, располагающиеся на грибообразных отростках. Шипик состоит из тонкой ножки, окончание которой утолщено в 3-5 раз, образуя головку шипика. На головке размещается один или несколько обычных синаптических контактов. В некоторых отделах мозга на шипиках расположено подавляющее большинство синапсов.
1.2. Процессы на мембране
Мембрана содержит пассивные и активные участки. Пассивная мембрана обладает способностью проводить отклонения потенциала в какой-либо точке от равновесного значения к другим точкам с затуханием, пропорциональным расстоянию. В активной мембране распространяется волна регенерирующего, возбуждения за счет вольт-зависимого изменения проводимости ионных каналов.
В спокойном состоянии мембраны среда внутри клетки имеет некоторый «потенциал покоя» (примерно 70 мВ) относительно внешней среды. Эта разность потенциалов определяется разной концентрацией ионов ряда элементов, главным образом ионов натрия и калия. В покое концентрация ионов калия на порядок ниже, чем с внутренней стороны (внутри нервной клетки или волокна).
Внутренняя поверхность мембраны заряжена отрицательно по отношению к ее наружной поверхности. Наблюдаемая разность концентраций ионов и потенциалов по обе стороны мембраны обусловлена процессами клеточного обмена - неким гипотетическим «метаболическим насосом», выкачивающим часть ионов натрия наружу и накачивающим часть ионов калия внутрь клетки.
Внутриклеточный потенциал может повышаться (деполяризация) или еще более понижаться (гиперполяризация).
Изменение потенциала в сторону деполяризации называют также возбуждающим постсинаптическим потенциалом (ВПСП), а в сторону гиперполяризации - тормозным постсинаптическим потенциалом (ТПСП). Соответственно все факторы, включая действие синапсов, называют возбуждающими или тормозными в зависимости от того, повышают или понижают они потенциал с внутренней стороны мембраны.
Возбуждающее воздействие, вызывающее повышение внутриклеточного потенциала (деполяризацию), не приведет к активному возбуждению мембраны, если не достигнут уровень возбуждения, т. е. изменение потенциала было меньше порогового. При этом порог по потенциалу зависит от формы возбуждающего импульса: чем положе восходящий фронт импульса, тем выше порог. Подпороговые изменения потенциала сопровождаются трансмембранным током, образованным емкостным током и током утечки.
При подпороговых процессах проницаемость мембраны для ионов существенно не меняется, поэтому и процессы относят к пассивным.
Активный процесс начинается с того момента, когда внутриклеточный потенциал превысил уровень порогового возбуждения. С этого момента резко увеличивается проницаемость мембраны для ионов натрия. Проходя снаружи через мембрану внутрь клетки (волокна), ионы натрия еще больше повышают внутренний потенциал, повышение потенциала увеличивает проницаемость для ионов натрия. Процесс нарастает лавинообразно до тех пор, пока разность потенциалов между внутренней и внешней средами не достигнет противоположного знака, затем ток натрия убывает. Зато постепенно увеличивается проницаемость мембраны для ионов калия. Калиевый ток течет навстречу натриевому. Он также достигает максимума и затем спадает. За счет тока натрия этот потенциал достигает своего максимума, а ток калия вновь сводит потенциал мембраны к значению, свойственному потенциалу покоя.
1.2.1. Активный транспорт ионов через мембрану
По современным представлениям в нейроне исходно существуют различные концентрации ионов калия внутри и снаружи клетки. Потенциал равновесия «запирает» выход ионов калия из клетки. Однако отрицательный потенциал создает условия движения в клетку ионов натрия из внешней среды, что в свою очередь снижает уровень отрицательного мембранного потенциала, облегчая выход калия под влиянием разницы концентраций. Постепенно концентрации ионов калия и натрия внутри и снаружи будут выравниваться. Для того чтобы поддержать эту разницу концентраций, должен существовать механизм вывода ионов натрия из клетки и переноса ионов калия в клетку.
Этот механизм называют натрий-калиевым насосом. Внутриклеточный натрий на внутренней стороне мембраны соединяется с веществом - носителем В. Комплекс NaB электронейтрален и благодаря диффузии выходит наружу, где расщепляется. Освободившийся носитель В под влиянием энзима переходит в форму С и присоединяет ион калия К+. Комплекс КС диффундирует внутрь клетки и отдает ион калия. Переносчик С при участии АТФ переходит в форму В с использованием внутренней энергии клетки (рис. 1.4), и цикл повторяется.
1.2.2. Элекгротонический потенциал.
Фиксация напряжения
При подаче импульса тока через введенный в тело нейрона микроэлектрод можно обнаружить, что потенциал нейрона, регистрируемый вторым микроэлектродом, изменяется. Нейрон деполяризуется при инъекции положительных зарядов д гиперполяризуется при введении отрицательных. Схематически мембрану нейрона можно представить как параллельно подключенные емкость и сопротивление. Емкость образована диэлектриком - фосфолипидами мембраны. Сопротивление создают каналы ионной проводимости. При достижении мембраной потенциала покоя, определяющегося соотношением потенциалов равновесия разных ионов, суммарный ток равен нулю.
При подаче импульса тока сначала он течет через емкость (емкостный ток). Падение напряжения на сопротивлении при этом равно нулю. По мере зарядки конденсатора все большая доля тока течет через сопротивление.
После зарядки конденсатора ток течет через мембрану, создавая на ней падение напряжения (рис. 1.5).
1.2.3. Потенциал действия
Методом фиксации напряжения было показано, что мембрана нейрона имеет быстро инактивирующуюся натриевую проводимость и устойчиво сохраняющуюся калиевую. Если не учитывать влияние емкостей, то нейрон можно представить схемой, состоящей из двух батарей и двух переменных сопротивлений. Одна батарея определяется потенциалом равновесия для калия (-75 мВ). Переменное сопротивление воспроизводит изменение калиевой проводимости. Другая батарея определяется потенциалом равновесия по натрию (+55 мВ) и переменным сопротивлением, воспроизводящим натриевую проводимость. Эти батареи и сопротивления образуют цепь, формирующую потенциал действия. При этом падение напряжения на мембране определяется соотношением проводимостей (рис. 1.6).
1.2.4. Суммация электротонических потенциалов
Электрические свойства мембраны нейрона характеризуются двумя константами: постоянной времени и постоянной длины. Постоянная времени мембраны определяет временную суммацию электротонических потенциалов: при поступлении на один и тот же участок мембраны двух электротонических потенциалов они суммируются на интервале времени, пропорциональном постоянной времени. Постоянная длины мембраны определяет пространственную суммацию электротонических потенциалов: если два электротонических потенциала одновременно поступают на два соседних участка пассивной мембраны, то к собственному потенциалу каждого участка добавляется часть потенциала соседнего участка, пропорциональная расстоянию между участками, деленному на постоянную длины мембраны.
Когда электротонические потенциалы возникают в разных участках мембраны в разные моменты времени, то их суммация определяется как постоянной времени, так и постоянной длины мембраны - имеет место так называемая пространственно-временная суммация.
Клетки с большими ^значениями этих констант более эффективно суммируют сигналы. Суммация позволяет мембранному потенциалу достичь критического значения генерации потенциала действия даже тогда, когда отдельные электротонические потенциалы этого уровня не достигают. Электротонические потенциалы определяются пассивными свойствами мембраны - емкостью и сопротивлением. Однако источником электротонических потенциалов в пассивных участках мембраны могут быть происходящие в прилежащих участках изменения потенциала, вызванные изменением ионной проводимости. Так, потенциал действия, генерируемый вследствие открывания натриевых каналов, создает в прилегающих участках, еще не охваченных процессом открытия натриевых каналов, деполяризационный сдвиг, определяемый константой длины мембраны. Аналогичным образом постоянная времени и постоянная длины вносят свой вклад в суммацию электротонических потенциалов, вызванных другими источниками: постсинаптическими потенциалами и пейсме- керными потенциалами.
1.2.5. Пейсмекерные потенциалы
В ряде нейронов даже после полной изоляции их сомы (ампутации дендритов и аксонов) при внутриклеточной регистрации наблюдаются колебания потенциала амплитудой 10-20 мВ и частотой от 1,0 до 2,5 Гц, получившие название пейсмекерных потенциалов. Эти потенциалы могут быть регулярными или образовывать отдельные веретена, в которых амплитуда потенциалов постепенно возрастает, достигает максимума и падает. Сравнение пейсмекерных потенциалов нейрона до и после изоляции от других нервных клеток указывает на то, что источником колебаний являются эндогенные процессы, поскольку тип активности сохраняется и в полностью изолированном нейроне. Если пейсмекерные потенциалы достигают порога генерации потенциала действия, то они запускают их генерацию.
Таким образом, благодаря наличию пейсмекерных потенциалов нейрон становится активным генератором. Более того, поскольку определенный тип нейрона и при полной изоляции сохраняет свой тип пейсмекерной активности, эта активность является генетически заданной. Вместе с тем пейсмекерный потенциал очень чувствителен к изменениям уровня мембранного потенциала: небольшая деполяризация клетки током до 1 нА достаточна для того, чтобы увеличить частоту и амплитуду пейсмекерных потенциалов, а гиперполяризация в тех же пределах ведет к замедлению пейсмекерных колебаний, их урежению, снижению амплитуды и, наконец, полному подавлению.
Анализируя пейсмекерные потенциалы, можно заметить, что при возрастании их амплитуды деполяризационная волна становится более острой. Это объясняется тем, что к пейсме- керному потенциалу присоединяется локальный потенциал, не достигающий порога развития лавинообразного процесса, связанного с генерацией потенциала действия.
Чувствительность пейсмекерных потенциалов к сдвигам мембранного потенциала делает их легко управляемыми внешними влияниями.
У ряда нейронов можно обнаружить несколько локусов пейсмекерных колебаний с разными свойствами и разным вкладом в механизмы генерации потенциалов действия. Источником пейсмекерных колебаний, по-видимому, являются электрогенные эффекты активного транспорта ионов. Переходы от деполяризации к гиперполяризации, образующие пейсмекерную волну, не сопровождаются изменением сопротивления мембраны. Активный транспорт ионов натрия ограничивается при развитии гиперполяризации. Активный транспорт ионов хлора ограничивается развитием деполяризации. Таким образом, пейсмекерный потенциал имеет зону оптимума, ограниченную сверху определенной деполяризацией, а снизу - определенной гиперполяризацией нейрона.
1.2.6. Электрический синапс
Электрический синапс образован срастанием мембран пре- синаптического и постсинаптического нейронов. Ток, инъецируемый в один нейрон через каналы, связывающие обе мембраны, затекает в постсинаптический нейрон пассивно и регистрируется на его мембране в виде электротонического потенциала.
Если один (прбсинаптический) нейрон генерирует потенциал действия, то у второго (постсинаптического) обнаруживается электротонический потенциал, в искаженной форме воспроизводящий за счет пассивных свойств мембраны потенциал действия пресинаптической мембраны в виде дифазного потенциала. При суммации дифазных потенциалов постсинаптическии нейрон сам генерирует потенциал действия, распространяющийся по аксону.
1.2.7. Квантовый механизм химического синапса
Электрический синапс содержит каналы, непосредственно связывающие пре- и постсинаптическую мембраны и обеспечивающие электротоническую связь между нейронами.
В отличие от электрического в химическом синапсе пре- и постсинаптическая мембраны разделены, и электротонический потенциал не принимает участия в связи нейронов. Связь реализуется посредством химических передатчиков - медиаторов. Медиатор находится в специальных гранулах - везикулах, находящихся в пресинаптической терминали. Под действием прибывшего в пресинаптическую терминаль импульса происходит присоединение везикулы к пресинаптической мембране, и в си- наптическую щель в сторону постсинаптической мембраны выбрасывается порция - квант медиатора. При этом медиатор действует на рецептивный белок постсинаптической мембраны. Этот рецептивный белок управляет каналами ионной проводимости. В зависимости от того, в отношении каких ионов в постсинаптической клетке регистрируется миниатюрный постсинаптическии потенциал (0,5-1 мВ) и как изменяется проводимость, спонтанно выделяются кванты медиатора, образуя в постсинаптичес- ком нейроне последовательность миниатюрных потенциалов постоянной амплитуды со случайными интервалами между ними. Миниатюрные потенциалы исчезают, если будут блокированы рецепторы, специфичные в отношении данного медиатора. В регуляции выделения медиатора принимает участие кальций. При деполяризации кальций входит в пресинаптическую часть синапса и перемещается к участкам, где везикулы присоединяются к участкам пресинаптической мембраны. Кальций облегчает слияние мембраны везикулы с мембраной терминали, в результате чего происходит выделение кванта медиатора.
Поступающий к пресинаптическому окончанию потенциал действия деполяризует термина ли, что приводит к открытию каналов кальциевой проводимости, которая в сильной степени зависит от уровня мембранного потенциала. Поступление кальция приводит к массированному выходу медиатора сразу из большого числа синаптических пузырьков. Суммация нескольких выделившихся в ответ на один потенциал действия квантов медиатора образует элементарный постсинаптический потенциал.
1.2.8. Транссинаптическая индукция энзимов
При действии медиатора на рецептор возникает комплекс, в котором белок обладает специфической конфигурацией. Такой белок может выполнять функцию индуктора: транс- портируясь к ядру клетки и связываясь с репрессором, он включает механизм транскрипции в определенном оперо- не, обеспечивая синтез группы функционально связанных энзимов. Энзимы, поступая к участкам синтеза медиатора, усиливают в качестве катализаторов синтез медиатора в пресинаптической терминали нейрона. Таким образом, реализуется схема медиатор ->- трансформация рецептивного белка ->- включение оперона на молекуле ДНК -*■ ->- транскрипция как синтез и-РНК ->- синтез энзимов -> ->- транспорт энзимов по аксону к пресинаптическому окончанию -* усиление синтеза медиатора в пресинапсе (рис. 1.7). Такой процесс носит название транссинапти- ческой индукции энзимов.
Наиболее подробно транссинаптическая индукция энзимов изучена на адренергических нейронах шейного узла, синтезирующих в термина лях своих аксонов медиатор норадреналин (или иначе, норэпинефрин). Нейроны этого узла, обладая мускориновыми и никотиновыми рецепторами, чувствительны к ацетилхолину. При присоединении ацетилхолина к никотиновым рецепторам через сутки в нейроне увеличивается содержание ферментов, выполняющих функцию катализаторов при синтезе норэпине- фрина. Синтезируемые в теле нейрона энзимы транспортируются по аксону к пресинаптическим окончаниям, где происходит повышенный синтез норэпинефрина. Вызванное действием ацетилхолина на никотиновые рецепторы усиление синтеза энзимов опосредовано процессом транскрипции с участием и-РНК. Это доказывается тем, что антибиотик актиномицин-D, препятствующий
Рис. 1.7. Транс-синаптическая индукция энзимов образованию молекулы и-РНК на матрице ДНК, включает транссинаптическую индукцию энзимов, катализирующих синтез норэпинефрина. Следует подчеркнуть, что индукция энзимов, ответственных за синтез норэпинефрина, происходит только в том случае, если ацетилхолин действует на никотиновые рецепторы. Если эти рецепторы блокировать, то индукции ответственных за синтез норэпинефрина энзимов не происходит.
Таким образом, аналогичный ферментативной адаптации механизм включен в регуляцию эффективности работы нейрона в звене синтеза медиатора.
1.3. Нейронные сети
Различают два крайних типа нейронных сетей: нелокальные нейронные сети и локальные. В нелокальных сетях нейрон выступает как структурная единица переработки информации. Локальные цепи принципиально отличаются от нелокальных тем, что в них единицей обработки информации является не весь нейрон, а отдельный участок его сомы, дендрита или аксона . Локальной называется цепь, образованная локальными участками одного или нескольких соседних нейронов, так что изменения в каждом из участков не затрагивают весь нейрон как целое.
Информационные свойства отдельного нейрона или его части существенно зависят от процессов, происходящих в дендритных и аксонных образованиях, и свойств его мембраны. Мембрана дендритов нейронов многих типов может быть возбудимой, способной проводить активное возбуждение. Морфологические данные говорят о наличии геометрически неоднородных участков в системах ветвлений дендритов и аксонов: расширений, узлов ветвления.
шипиков, касательных контактов . Такие неоднородности участвуют в процессах переработки информации нейроном, являются элементарными звеньями нейрона, преобразующими информацию. Вблизи неоднородностей меняется скорость проведения возбуждения, амплитуда активного и пассивного процессов, форма импульса возбуждения . В зависимости от вида неоднородности выходной сигнал будет представлять собой результат той или иной операции .
Информационные свойства локальных цепей определяются структурно-функциональными особенностями входящих в состав цепи участков нейронов, а также определяются характером латеральных возбуждающих и тормозных взаимодействий между нейронами сети.
1.3.1. Локальные цепи
Можно условно выделить локальные дендритные, локальные аксонные и смешанные локальные цепи. Локальные дендритные цепи - это такие образования, в которых имеются дендро-дендритные контакты как в структуре одного нейрона, так и между дендритами соседних нейронов.
Локальные аксонные цепи образуют аксо-аксональные касательные контакты и анастомозные соединения аксонов разных нейронов. Смешанными локальными цепями назовем структуры из аксонных и дендритных ветвлений, в которых имеются контакты разных типов: дендро-ден- ритные, аксо-аксональные, ак- со-дендритные, дендро-сомати- ческие и т. д. (рис. 1.8). Ден- дриты и аксоны могут иметь структурно-функциональные неоднородности, способствующие расширению набора операций, выполняемых в локальных цепях.
Простейшим случаем локальных цепей являются локальные аксонные цепи с аксо-аксональными синапсами, когда под влиянием импульса, проходящего по аксону одного нейрона, срабатывает одна из аксонных термина- леи другого - невозбужденного нейрона, не влияя на аксонные ветви этого нейрона. Сома нейрона выполняет в этом случае функцию питания ветвей, а информационные процессы в ветвях развиваются независимо.
Более сложные локальные цепи образуются в результате дендро-дендритных контактов разных нейронов.
В этом случае происходит локальная обработка информации во множестве точек локального взаимодействия дендритов. Следует учитывать, что при локальном взаимодействии на близких расстояниях генерация импульса не обязательна. Достаточно, чтобы один синапс оказал влияние в близком соседстве от другого и его эффективность изменилась за счет электротонического эффекта, связанного с входом кальция в рассматриваемый локальный участок мембраны.
Возможность образования отдельными участками мембраны нейрона независимых параллельно работающих локальных цепей означает, что единицей обработки является не нейрон как целое, а его отдельные локусы. В связи с этим функция нелокальных сетей является вторичной по отношению к информации, предварительно обработанной в локальных нейронных цепях. При помощи аксонных систем сигналы нейронной сети передаются затем на значительные расстояния.
1.3.2. Организация нейронных сетей мозга
Под нейронной сетью понимается нелокальная нейронная сеть из нервных клеток, между которыми заданы перекрестные (латеральные) возбуждающие и тормозные взаимодействия, в результате которых нейронные сети оказываются приспособленными для выполнения разнообразных информационных преобразований.
Выделяют несколько типов тормозных латеральных взаимодействий между нейронами сети: непосредственное латеральное торможение соседних нейронов через тормозные коллатерали; коллатеральное торможение через вставочный тормозный нейрон; возвратное торможение, заключающееся в том, что выходной сигнал нейрона возвращается к нему через тормозную коллатераль аксона и приводит к торможению активности нейрона. Возвратное торможение может осуществляться по типу непосредственного или коллатерального торможения. Пресинаптическое заключается во влиянии синаптическпх контактов одного нейрона на синаптические контакты другого, уменьшающем активность этих контактов.
Торможение между нейронами может осуществляться как через химические, так и через электрические синапсы.
Бывает изотропное латеральное торможение, равномерно распределенное между нейронами, и анизотропное латеральное торможение, когда в одном направлении торможение больше или меньше, чем в других.
Рассмотрим организацию нейронных сетей различных отделов мозга.
Новая кора. Основной структурно-функциональной единицей (элементарной нейронной сетью) новой коры служит колонка - вертикально расположенная группа клеток, связанных между собой преимущественно вертикальными связями. Кортикальная колонка нейронов образует модуль обработки информации, выделяющий один определенный признак сигнала. Расположение нейронов в виде множества колонок позволяет выделять несколько переменных в локальном участке топографического коркового представительства. Таким образом, с помощью вертикальных колонок в трехмерную структуру новой коры встраивается аппарат многомерного отображения внешней среды.
Так, форма кожной поверхности тела представлена в виде карты на постцентральной соматосенсорной коре.
Каждый участок кожи представлен группой локальных колонок, каждая из которых содержит нейроны, отвечающие на данную модальность стимула в данном участке кожи. Другим примером колончатой организации коры мозга является представительство вибрисс крысы в коре.
Каждая вибрисса представлена колонкой клеток, которые возбуждаются только при движении данной вибриссы .
Различают следующие типы корковых колонок: 1)ми- кроколонки - в них представлены градации выделяемого признака; 2) гиперколонки, каждая из которых содержит набор микроколонок (в гиперколонке представлен набор градаций данного признака); 3) макроколонки, каждая из которых содержит набор гиперколонок (в каждой макроколонке представлен набор признаков в данном топографически локальном участке коры больших полуша- рий, представляющем участок рецептивной поверхности).
Гиперколонка представляет собой корковую часть локального анализатора определенного свойства.
Макроколонка представляет набор таких локальных анализаторов.
Тангенциальные волокна серого вещества, распространяющиеся на 1-2 мм, по-видимому, создают латеральное торможение между гиперколонками, выделяющими одни и те же свойства в разных макроколонках. Следовательно, обработка информации о разных свойствах сигнала в разных точках пространства происходит в параллельных каналах.
Вверх от IV слоя идут нейроны отображения сигнала, нейроны V и VI слоев являются исполнительными элементами. Нейрон - элемент микроколонки является нейроном-детектором. Совокупность детекторов одного класса в гиперколонке одного топографического локуса образует квазирецептивную поверхность. Зрительная кора. Зрительная кора приматов образована вертикальными колонками нейронов. В этих колонках представлены доминантные глаза, а также колонки нейронов, чувствительных к разной ориентации участка линии, находящегося в пределах топографически локального участка проекции сетчатки. Отрезок прямой линии в поле зрения возбуждает группу макроколонок, в которой, в свою очередь, возбуждаются гиперколонки определенного класса, а именно гиперколонки, выделяющие ориентацию линии. В этих гиперколонках будут возбуждены только те микроколонки, оптимальная чувствительность к ориентации линии у которых совпадает с ориентацией линии .
Простые клетки зрительной коры.
В соответствии с функциональными свойствами в зрительной коре кошки выделяют два основных класса нейронов: клетки с простыми рецептивными полями (простые) и клетки со сложными рецептивными полями (сложные) . Область рецептивного поля, в которой стационарный стимул вызывает разряд, называется полем разряда. Поле разряда образует только часть рецептивного поля. Другие участки рецептивного поля оказывают усиливающее или ослабляющее действие. Простые клетки оптимально отвечают при определенной ориентации линии в их рецептивном поле . К возбуждающему рецептивному полю клеток примыкают тормозные зоны.
Простые клетки бывают двух типов. Одни построены из концентрических нейронов наружного коленчатого тела с возбуждающими центрами, другие из нейронов с тормозящими центрами. Тормозные зоны построены из тех же элементов, что и возбуждающие. Таким образом, одни клетки отвечают на светлые линии при темном фоне, другие - на темные линии при светлом фоне. Клетки наружного коленчатого тела в отличие от простых клеток коры имеют концентрические, а не сильно сжатые эллиптические рецептивные поля. Возбуждающее и тормозное поля простой рецептивной клетки не совпадают по центру.
Оптимальная ориентация рецептивного поля перпендикулярна линии, соединяющей центры возбуждающего и тормозного полей .
Сложные клетки зрительной коры. В отличие от простых клеток сложные клетки зрительной коры характеризуются большими рецептивными полями.
Линия оптимальной ориентации, расположенная в рецептивном поле такой клетки, вызывает ответ на включение и выключение. Сложная клетка отвечает серией разрядов при движении линии в ее рецептивном поле .
Рецептивное поле сложных клеток построено из клеток латерального коленчатого тела с возбуждающими и тормозными центрами. Это относится к возбуждающей и тормозной зонам .
Сложные клетки, так же как и простые, имеют смещенные по центру перекрывающиеся рецептивные поля.
Оптимальный наклон линии перпендикулярен линии, соединяющей центры полей. Сложные клетки возбуждаются как при действии светлых, так и темных полос. Кроме того, реакция на свет и темноту сразу же компенсируется сигналами противоположного знака. Таким образом, основное различие между простыми и сложными клетками обусловлено разным типом входов. Простые клетки имеют входы либо от ОН-, либо от ОФФ-нейронов. Сложные клетки получают сигналы и от ОН-, и от ОФФ-нейронов. При действии как темных, так и светлых полос имеет место сильное торможение по краям рецептивного поля .
Ретинотопическая проекция в зрительной коре. В зрительной коре обезьяны существует топографическая карта зрительного поля. Эта карта характеризуется «фактором увеличения», который показывает, какой размер участка представляет данную область зрительного поля. Фактор увеличения измеряется в мм/град. Представленный как функция эксцентриси- тета фактор увеличения показывает, что область фовеа представлена в зрительной коре большими участками, чем периферические участки поля зрения. Максимальное значение фактора увеличения равно 6 мм/град.
Острота зрения измеряется минимальным углом различения точек на сетчатке. Для фовеа острота зрения равна 0,67 мин. На зрительной коре этому разрешению сигналов соответствует расстояние 67 мкм. При измерении размеров рецептивного поля нейрона зрительной коры оказалось, что гиперколонки (полный набор колонок нейронов с разной оптимальной ориентацией) расположены в зрительной коре на расстоянии 1-2 мм. Локальный образ в зрительной коре возникает в пределах одной гиперколонки. Различие двух стимулов на уровне коры определяется размером рецептивного поля .
Развитие нейронов зрительной системы. Нейроны латерального коленчатого тела релейного ядра зрительной системы формируются в течение недели около 40-го дня эмбрионального развития. Синаптический аппарат начинает складываться 20 дней спустя. Быстрое увеличение размеров и клеточных тел происходит в первый месяц пост- натальной жизни. В условиях зрительной депривации этот процесс нарушается. В позднем пренатальном и раннем постнатальном периоде обнаруживают уже слоистую структуру. Основными признаками незрелости нейронов являются: наличие большого числа похожих на волоски выростов на дендритах и утолщений в виде колбочек, связанных с процессом роста .
В конце первого месяца постнатальной жизни волоски и утолщения на дендритах исчезают. Степень морфологических изменений у разных типов нейронов различна.
Мультиполярные нейроны характеризуются несколькими основными дендритами, берущими начало от сомы нейрона. Такие нейроны встречаются во всех слоях с первого до шестого. Однако имеет место градиент в степени их зрелости. Так, когда первый слой содержит нейроны с развитым дендритным деревом уже без волосков и утолщений, дендриты мультиполярных нейронов пятого и шестого слоев еще имеют волоски и утолщения, а их дендритные ветви развиты слабо.
Биполярные нейроны имеют или два основных дендрита, или три, в последнем случае у них форма треугольника. Биполярные нейроны обладают большей степенью зрелости, чем мультиполярные нейроны.
Нейроны с дендритом, напоминающим аксон, характеризуются тонкими цилиндрическими дендритами, длина которых увеличивается в процессе развития. Нейроны с веерообразными дендритами приобретают черты взрослого состояния еще в эмбриональной стадии.
Общая схема созревания нейронов наружного коленчатого тела включает: 1) появление участков роста в виде колбочек (ранний пренатальный период); 2) появление тонких волосков на дендритах (поздний пренатальный период); 3) редукция участков роста и исчезновение волосков (несколько дней постнатального периода).
Эффективность зрительной системы обезьяны, проявляющаяся сразу же после ее рождения, является следствием пренатальной дифференциации нейронов наружного коленчатого тела. Позже следует лишь созревание с формированием шипиков и дистальных участков дендритных ветвей. Полное созревание дендритной системы завершается через четыре недели после рождения .
Нейроны зрительной коры формируются под влиянием факторов внешней среды . При этом нейроны IV слоя коры, получающие афференты от наружного коленчатого тела, настроены селективно на определенный наклон линии генетически . Что касается нейронов верхних слоев коры, образующих колонку, то они приобретают селективность в зависимости от того, какие нейроны IV слоя возбуждаются внешними сигналами. Так, в «вертикальной среде» они образуют связи с детекторами вертикальных линий IV слоя, а в «горизонтальной среде» - с детекторами горизонтальных линий IV слоя. В результате в ограниченной среде формируются только такие колонки, которые имеют характеристики возбуждающихся нейронов IV слоя .
Исследования поглощения диоксиглюкозы нейронами зрительной коры в условиях стимуляции глаза кошки вертикальными или горизонтальными решетками показали, что клетки, обладающие общей ориентационной избирательностью, образуют на поверхности коры параллельные полосы, ортогональные представительству вертикального меридиана. Нейроны, характеризующиеся общей селективной ориентацией, образуют колонку. Аналогичным образом организована система представительства доминантного глаза .
Если котята воспитывались в «горизонтальной» или «вертикальной» среде, то поглощение глюкозы было раз- личным при вертикальной и горизонтальной стимуляции.
При совпадении ориентации линий в опыте животного с ориентацией тест-стимула обнаруживались более широкие полосы, образованные нейронами одной ориентации.
При несовпадении полосы были узкими и были образованы почти исключительно нейронами IV слоя коры, куда поступают афференты от наружного коленчатого тела
Таким образом, нейроны IV слоя обладают ориента- ционной селективностью, которая определяется генетически и не зависит от зрительного опыта. Нейроны же более высоких слоев коры приобретают ориентационную избирательность под влиянием зрительной стимуляции, которая исчезает в критический период развития .