Образование нативной конформации белка. Роль первичной структуры
Рис. 3.9. Третичная структура лактоглобулина - типичного а/р-белка (по PDB-200I) (Brownlow, S., Marais Cabral, J. H., Cooper, R., Flower, D. R., Yewdall, S. J., Polikarpov, I., North, A. C., Sawyer, L.: Structure, 5, p. 481. 1997)
Пространственная структура зависит не от длины полипептидной цепи, а от последовательности аминокислотных остатков, специфичной для каждого белка, а также от боковых радикалов, свойственных соответствующим аминокислотам. Пространственную трехмерную структуру или конформацию белковых макромолекул образуют в первую очередь водородные связи, а также гидрофобные взаимодействия между неполярными боковыми радикалами аминокислот. Водородные связи играют огромную роль в формировании и поддержании пространственной структуры белковой макромолекулы. Водородная связь образуется между двумя электроотрицательными атомами посредством протона водорода, ковалентно связанного с одним из этих атомов. Когда единственный электрон атома водорода участвует в образовании электронной пары, то протон притягивается соседним атомом, образуя водородную связь. Обязательным условием образования водородной связи является наличие хотя бы одной свободной пары электронов у электроотрицательного атома. Что касается гидрофобных взаимодействий, то они возникают в результате контакта между неполярными радикалами, неспособными разорвать водородные связи между молекулами воды, которая вытесняется на поверхность белковой глобулы. По мере синтеза белка неполярные химические группировки собираются внутри глобулы, а полярные вытесняются на ее поверхность. Таким образом, белковая молекула может быть нейтральной, заряженной положительно или же отрицательно в зависимости от pH растворителя и ионогенных групп в белке. К слабым взаимодействиям относят также ионные связи и ван-дер-ваальсовы взаимодействия. Кроме того, конформация белков поддерживается ковалентными связями S-S, образующимися между двумя остатками цистеина. В результате гидрофобных и гидрофильных взаимодействий молекула белка спонтанно принимает одну или несколько наиболее термодинамически выгодных конформаций, причем, если в результате каких-либо внешних воздействий нативная конформация нарушается, возможно полное или почти полное ее восстановление. Впервые это показал К. Анфинсен на примере каталитически активного белка рибонуклеазы. Оказалось, что при воздействии мочевиной или p-меркаптоэтанолом происходит изменение ее конформации и, как следствие, резкое снижение каталитической активности. Удаление мочевины приводит к переходу конформации белка в исходное состояние, и каталитическая активность восстанавливается.
Таким образом, конформация белков представляет собой трехмерную структуру, причем в результате ее образования многие атомы, находящиеся на удаленных участках полипептидной цепи, сближаются и, воздействуя друг на друга, приобретают новые свойства, отсутствующие у индивидуальных аминокислот или небольших полипептидов. Это так называемая третичная структура , характеризующаяся ориентацией полипептидных цепей в пространстве (рис. 3.9). Третичная структура глобулярных и фибриллярных белков существенно отличается друг от друга. Принято форму белковой молекулы характеризовать таким показателем, как степень асимметрии (отношение длинной оси молекулы к короткой). У глобулярных белков степень асимметрии равна 3-5, что касается фибриллярных белков, то эта величина у них гораздо больше (от 80 до 150).
Каким же образом первичная и вторичная развернутые структуры преобразуются в свернутую, весьма стабильную форму? Расчеты показывают, что число теоретически возможных комбинаций образования трехмерных структур белков неизмеримо больше, чем реально существующих в природе. По-видимому, основным фактором конформационной стабильности являются энергетически наиболее выгодные формы.
Гипотеза расплавленной глобулы. Одним из способов изучения сворачивания полипептидной цепи в трехмерную структуру является денатурация и последующая рснатурация белковой молекулы.
Опыты К. Анфинсена с рибонуклеазой однозначно показывают возможность сборки именно той пространственной структуры, которая была нарушена в результате денатурации (рис. 3.10).
В данном случае восстановление нативной конформации не требует наличия никаких дополнительных структур. Какие же модели свертывания полипептидной цепи в соответствующую конформацию являются наиболее вероятными? Одной из распространенных гипотез самоорганизации белка является гипотеза расплавленной глобулы. В рамках этой концепции выделяют несколько этапов самосборки белков.
- 1. В развернутой полипептидной цепи с помощью водородных связей и гидрофобных взаимодействий образуются отдельные участки вторичной структуры, служащие как бы затравками для формирования полных вторичных и супервторичных структур.
- 2. Когда число этих участков достигает определенной пороговой величины, происходит переориентация боковых радикалов и переход полипептидной цепи в новую более компактную форму, причем число нековалентных связей

Рис. 3.10.
значительно увеличивается. Характерной особенностью этой стадии является образование специфических контактов между атомами, находящимися на удаленных участках полипептидной цепи, но оказавшихся сближенными в результате образования третичной структуры.
3. На последнем этапе формируется нативная конформация белковой молекулы, связанная с замыканием дисульфидных связей и окончательной стабилизацией белковой конформации. Не исключена также неспецифическая агрегация частично сверну
тых полипсптидных цепей, что можно квалифицировать как ошибки образования нативных белков. Частично свернутая полипептидная цепь (этап 2) называется расплавленной глобулой, а этап 3 является самым медленным при образовании зрелого белка.
На рис. 3.11 показан вариант образования белковой макромолекулы, кодируемой одним геном. Известно, однако, что ряд белков, имеющих домен-

Рис. 3.11.
(по Н. К. Наградовой) ную структуру, формируется в результате дубликации генов, и образование контактов между отдельными доменами требует дополнительных усилий. Оказалось, что в клетках имеются специальные механизмы регуляции процессов сворачивания новосинтезиро- ванных белков. В настоящее время обнаружено два фермента, участвующих в реализации этих механизмов. Одной из медленных реакций третьего этапа сворачивания полипептидных цепей является *

Рис. 3.12.
Кроме того, в клетках имеется ряд каталитически неактивных белков, которые тем не менее вносят большой вклад в образование пространственных структур белков. Это так называемые шапероны и шаперонины (рис. 3.12). Один из первооткрывателей молекулярных шаперонов, Л. Эллис, называет их функциональным классом не связанных друг с другом семейств белков, которые помогают правильной нековалентной сборке других полипептидсодержа- ших структур in vivo, но не входят в состав собираемых структур и не участвуют в реализации их нормальных физиологических функций.
Шапероны помогают правильной сборке трехмерной белковой конформации путем образования обратимых нековалентных комплексов с частично свернутой полипептидной цепью, одновременно ингибируя неправильно образованные связи, ведущие к формированию функционально неактивных белковых структур. В перечень функций, свойственных шаперонам, входит защита расплавленных глобул от агрегации, а также перенос новосинтезированных белков в различные локусы клеток. Шапероны преимущественно являются белками теплового шока, синтез которых резко усиливается при стрессовом температурном воздействии, поэтому их называют еще hsp (heat shock proteins). Семейства этих белков найдены в микробных, растительных и животных клетках. Классификация шаперонов основана на их молекулярной массе, которая варьирует от 10 до 90 kDa. В основном функции шаперонов и шаперо- нинов различаются, хотя и те, и другие являются белками-помошниками процессов образования трехмерной структуры белков. Шапероны удерживают новосинтсзированную полипептидную цепь в развернутом состоянии, не давая ей свернуться в отличную от нативной форму, а шаперонины обеспечивают условия для образования единственно правильной, нативной структуры белка (рис. 3.13).

Рис. 3.13.
Шапероны / связаны с нансцентной полипептидной цепью, сходящей с рибосомы. После образования полипептидной цепи и выхода ее из рибосомы шапероны соединяются с ней и препятствуют агрегации 2. После сворачивания в цитоплазме белки отделяются от шаперона и переходят на соответствующий шаперо- нин, где и происходит окончательное образование третичной структуры 3. При помощи цитозольного шаперона белки перемещаются к внешней мембране митохондрии, где митохондриальный шапе- рон протягивает их вовнутрь митохондрий и «передает» митохондриальному шаперонину, где и происходит сворачивание 4, а 5 аналогично 4 , но применительно к эндоплазматическому ретикулуму.
Биохимия - это наука о молекулярных основах жизни, занимается изучением молекул, химических реакций, процессов, протекающих в живых клетках организма. Подразделяется на:
статическую (строение и свойства биомолекул)
динамическую (химизм реакций)
специальные разделы (экологическая, биохимия микроорганизмов, клиническая)
Роль биохимии в решении основополагающих медицинских проблем
сохранение здоровья человека
выяснение причин различных заболеваний и изыскание путей их эффективного лечения.
Таким образом, любое недомогание, заболевание человека связано с нарушением строения и свойств метаболитов или биомолекул, также связано с изменениями биохимических реакций, протекающих в организме. Применение любых способов лечения, лекарственных препаратов также основывается на понимании и точном знании биохимизма их действия.
Белки, их строение и биологическая роль
Белки - это высокомолекулярные полипептиды, условная граница между белками и полипептидами обычно составляет 8000-10000 единиц молекулярной массы. Полипептиды - это полимерные соединения, имеющие более 10 остатков аминокислот в молекуле.
Пептиды - это соединения, состоящие из двух и более остатков аминокислот (до 10) В состав белков входят только L-аминокислоты.
Встречаются производные аминокислот, например, в состав коллагена входит гидроксипролин и гидроксилизин. В некоторых белках обнаруживается у -карбоксиглутамат. Нарушение карбоксилирования глутамата в протромбине может привести к кровотечению. Часто в белках встречается фосфосерин.
Незаменимые аминокислоты - это те, которые не синтезируются в организме или
синтезируются в недостаточном количестве или с малой скоростью.
Для человека незаменимыми являются 8 аминокислот: триптофан, фенилаланин,
метионин, лизин, валин, треонин, изолейцин, лейцин.
Биохимические функции аминокислот:
строительные блоки пептидов, полипептидов и белков,
биосинтез других аминокислот (из фенилаланина синтезируется тирозин, из метионина - цистеин)
биосинтез некоторых гормонов, например, окситацина, вазопрессина, инсулина
исходные продукты для образования глутатиона, креатина
глицин необходим для синтеза порфирина
р - аланин, валин, цистеин образуют КоА, триптофан - никотинамид, глутаминовая кислота - фолиевую кислоту
для биосинтеза нуклеотидов необходим глутамин, глицин, аспарагиновая кислота, они образуют пуриновые основания, глутамин и аспарагиновая кислота - пиримидиновые
11 аминокислот являются глюкогенными, то есть способны метаболизироваться в глюкозу и другие УВ
фенилаланин, тирозин, лейцин, лизин и триптофан принимают участие в биосинтезе некоторых липидов
10.образование мочевины, углекислоты и энергии в виде АТФ.
Строение белков. Первичная структура.
Под первичной структурой понимают последовательность аминокислот в цепи, они соединены между собой ковалентными пептидными связями. Полипептидная цепь начинается с остатка, имеющего свободную аминогруппу (N - конец) и завершается свободным СООН - концом.
К первичной структуре также относят взаимодействие между остатками цистеина с образованием дисульфидных связей.
Таким образом, первичная структура - это описание всех ковалентных связей в молекуле белка.
Пептидная связь отличается полярностью, что обусловлено тем, что связь между N и С частично носит характер двойной связи. Вращение затруднено и пептидная связь имеет жесткую структуру. Последовательность аминокислот генетически строго детерминирована, она определяет нативную природу белка и его функции в организме.
Вторичная структура
1951 г. - была расшифрована вторичная структура (тугозакрученная основная цепь полипептида, которая составляет внутреннюю часть стержня, боковые цепи направлены наружу, располагаясь по спирали) Все -С=О- N-H- группы оснований цепи связаны водордными связями.
Водородные связи делают а - спираль более устойчивой.
Другой тип вторичной структуры - это р - складчатый слой. Это параллельно лежащие полипептидные цепи, которые сшиты водородными связями. Возможно закручивание таких р - образований, что придает белку большую прочность.
Третий тип вторичной структуры характерен для коллагена. Каждая из трех полипептидных цепей предшественника коллагена (тропоколлаген) имеет форму спирали. Три такие спирализованные цепи закручиваются относительно друг друга, образуя тугую нить.
Специфика такого типа структуры обусловлена наличием водородных связей сугубо между остатками глицина, пролина и гидроксипролина, а также внутри- и межмолекулярных ковалентных поперечных связей.
МОДУЛЬ 1 СТРОЕНИЕ, СВОЙСТВА И ФУНКЦИИ БЕЛКОВ
МОДУЛЬ 1 СТРОЕНИЕ, СВОЙСТВА И ФУНКЦИИ БЕЛКОВ
Структура модуля | Темы |
Модульная единица 1 | 1.1. Структурная организация белков. Этапы формирования нативной конформации белков 1.2. Основы функционирования белков. Лекарства как лиганды, влияющие на функцию белков 1.3. Денатурация белков и возможность их спонтанной ренативации |
Модульная единица 2 | 1.4. Особенности строения и функционирования олигомерных белков на примере гемоглобина 1.5. Поддержание нативной конформации белков в условиях клетки 1.6. Многообразие белков. Семейства белков на примере иммуноглобулинов 1.7. Физико-химические свойства белков и методы их разделения |
Модульная единица 1 СТРУКТУРНАЯ ОРГАНИЗАЦИЯ МОНОМЕРНЫХ БЕЛКОВ И ОСНОВЫ ИХ ФУНКЦИОНИРОВАНИЯ
Цели изучения Уметь:
1. Использовать знания об особенностях структуры белков и зависимости функций белков от их структуры для понимания механизмов развития наследственных и приобретенных протеинопатий.
2. Объяснять механизмы лечебного действия некоторых лекарств как лигандов, взаимодействующих с белками и изменяющих их активность.
3. Использовать знания о строении и конформационной лабильности белков для понимания их структурно-функциональной неустойчивости и склонности к денатурации в изменяющихся условиях.
4. Объяснять применение денатурирующих агентов в качестве средств для стерилизации медицинского материала и инструментов, а также в качестве антисептиков.
Знать:
1. Уровни структурной организации белков.
2. Значение первичной структуры белков, определяющей их структурное и функциональное многообразие.
3. Механизм формирования в белках активного центра и его специфическое взаимодействие с лигандом, лежащее в основе функционирования белков.
4. Примеры влияния экзогенных лигандов (лекарств, токсинов, ядов) на конформацию и функциональную активность белков.
5. Причины и следствия денатурации белков, факторы, вызывающие денатурацию.
6. Примеры использования денатурирующих факторов в медицине в качестве антисептиков и средств для стерилизации медицинских инструментов.
ТЕМА 1.1. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ БЕЛКОВ. ЭТАПЫ ФОРМИРОВАНИЯ НАТИВНОЙ
КОНФОРМАЦИИ БЕЛКОВ
Белки - это полимерные молекулы, мономерами которых являются всего 20 α-аминокислот. Набор и порядок соединения аминокислот в белке определяется строением генов в ДНК индивидумов. Каждый белок в соответствии с его специфической структурой выполняет свойственную ему функцию. Набор белков данного организма определяет его фенотипические особенности, а также наличие наследственных болезней или предрасположенность к их развитию.
1. Аминокислоты, входящие в состав белков. Пептидная связь. Белки - полимеры, построенные из мономеров - 20 α-аминокислот, общая формула которых
Аминокислоты различаются по строению, размерам, физико-химическим свойствам радикалов, присоединенных к α-углеродному атому. Функциональные группы аминокислот определяют особенности свойств разных α-аминокислот. Встречающиеся в α-аминокислотах радикалы можно разделить на несколько групп:
Пролин, в отличие от других 19 мономеров белков, не аминокислота, а иминокислота, радикал в пролине связан как с α-углеродным атомом, так и с иминогруппой
 Аминокислоты различаются по растворимости в воде.
Это связано со способностью радикалов взаимодействовать с водой (гидратироваться).
Аминокислоты различаются по растворимости в воде.
Это связано со способностью радикалов взаимодействовать с водой (гидратироваться).
К гидрофильным относятся радикалы, содержащие анионные, катионные и полярные незаряженные функциональные группы.
К гидрофобным относятся радикалы, содержащие метильные группы, алифатические цепи или циклы.
2. Пептидные связи соединяют аминокислоты в пептиды. При синтезе пептида α-карбоксильная группа одной аминокислоты взаимодействует с α-аминогруппой другой аминокислоты с образованием пептидной связи:
Белки представляют собой полипептиды, т.е. линейные полимеры α-аминокислот, соединенных пептидной связью (рис. 1.1.)
 Рис. 1.1. Термины, используемые при описании строения пептидов
Рис. 1.1. Термины, используемые при описании строения пептидов
Мономеры аминокислот, входящих в состав полипептидов, называются аминокислотными остатками. Цепь повторяющихся групп -NH-CH-CO - образует пептидный остов. Аминокислотный остаток, имеющий свободную α-аминогруппу, называется N-концевым, а имеющий свободную α-карбоксильную группу - С-концевым. Пептиды записывают и читают с N-конца к С-концу.
Пептидная связь, образуемая иминогруппой пролина, отличается от других пептидных связей: у атома азота пептидной группы отсутствует водород,
вместо него имеется связь с радикалом, в результате одна сторона цикла включается в пептидный остов:
 Пептиды
различаются аминокислотным составом, количеством аминокислот и порядком
соединения аминокислот, например, Сер-Ала-Глу-Гис и Гис-Глу-Ала-Сер -
два разных пептида.
Пептиды
различаются аминокислотным составом, количеством аминокислот и порядком
соединения аминокислот, например, Сер-Ала-Глу-Гис и Гис-Глу-Ала-Сер -
два разных пептида.
Пептидные связи очень прочные, и для их химического неферментативного гидролиза требуются жесткие условия: анализируемый белок гидролизуют в концентрированной соляной кислоте при температуре около 110° в течение 24 часов. В живой клетке пептидные связи могут разрываться с помощью протеолитических ферментов, называемых протеазами или пептидгидролазами.
3. Первичная структура белков. Аминокислотные остатки в пептидных цепях разных белков чередуются не случайным образом, а расположены в определенном порядке. Линейная последовательность или порядок чередования аминокислотных остатков в полипептидной цепи называется первичной структурой белка.
Первичная структура каждого индивидуального белка закодирована в молекуле ДНК (в участке, называемом геном) и реализуется в ходе транскрипции (переписывания информации на мРНК) и трансляции (синтез первичной структуры белка). Следовательно, первичная структура белков индивидуального человека - наследственно передаваемая от родителей детям информация, определяющая особенности строения белков данного организма, от которых зависит функция имеющихся белков (рис. 1.2.).
 Рис. 1.2. Взаимосвязь между генотипом и конформацией белков, синтезирующихся в организме индивидума
Рис. 1.2. Взаимосвязь между генотипом и конформацией белков, синтезирующихся в организме индивидума
Каждый из примерно 100 000 индивидуальных белков в организме человека имеет уникальную первичную структуру. В молекулах одного типа белка (например, альбумина) одинаковое чередование аминокислотных остатков, что отличает альбумин от любого другого индивидуального белка.
Последовательность аминокислотных остатков в пептидной цепи можно рассматривать как форму записи информации. Эта информация определяет пространственную укладку линейной пептидной цепи в более компактную трехмерную структуру, называемую конформацией белка. Процесс формирования функционально активной конформации белка носит название фолдинг.
4. Конформация белков. Свободное вращение в пептидном остове возможно между атомом азота пептидной группы и соседним α-углеродным атомом, а также между α-углеродным атомом и углеродом карбонильной группы. Вследствие взаимодействия функциональных групп аминокислотных остатков первичная структура белков может приобретать более сложные пространственные структуры. В глобулярных белках различают два основных уровня укладки конформации пептидных цепей: вторичную и третичную структуры.
Вторичная структура белков - это пространственная структура, формирующаяся в результате образования водородных связей между функциональными группами -С=О и - NH- пептидного остова. При этом пептидная цепь может приобретать регулярные структуры двух типов: α-спирали и β-структуры.
В α-спирали водородные связи образуются между атомом кислорода карбонильной группы и водородом амидного азота 4-й от него аминокислоты; боковые цепи аминокислотных остатков
располагаются по периферии спирали, не участвуя в образовании вторичной структуры (рис. 1.3.).
Объемные радикалы или радикалы, несущие одинаковые заряды, препятствуют формированию α-спирали. Остаток пролина, имеющий кольцевую структуру, прерывает α-спираль, так как из-за отсутствия водорода у атома азота в пептидной цепи невозможно образовать водородную связь. Связь между азотом и α-углеродным атомом входит в состав цикла пролина, поэтому пептидный остов в этом месте приобретает изгиб.
β-Структура формируется между линейными областями пептидного остова одной полипептидной цепи, образуя при этом складчатые структуры. Полипептидные цепи или их части могут формировать параллельные или антипараллельные β-структуры. В первом случае N- и С-концы взаимодействующих пептидных цепей совпадают, а во втором - имеют противоположное направление (рис. 1.4).
Рис. 1.3. Вторичная структура белка - α-спираль
 Рис. 1.4. Параллельные и антипараллельные β-складчатые структуры
Рис. 1.4. Параллельные и антипараллельные β-складчатые структуры
β-структуры обозначены широкими стрелками: А - Антипараллельная β-структура. Б - Параллельные β-складчатые структуры
В некоторых белках β-структуры могут формироваться за счет образования водородных связей между атомами пептидного остова разных полипептидных цепей.
В белках также встречаются области с нерегулярной вторичной структурой, к которым относят изгибы, петли, повороты полипептидного остова. Они часто располагаются в местах, где меняется направление пептидной цепи, например, при формировании параллельной β-складчатой структуры.
По наличию α-спиралей и β-структур глобулярные белки могут быть разделены на четыре категории.
Рис. 1.5. Вторичная структура миоглобина (А) и β-цепи гемоглобина (Б), содержащие восемь α-спиралей
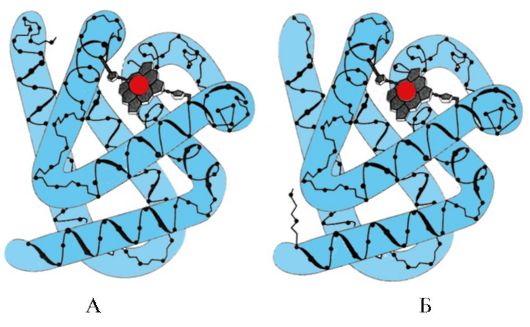
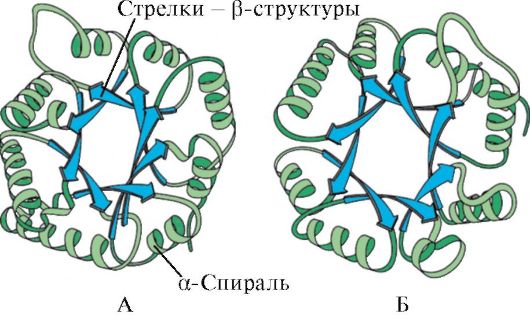 Рис. 1.6. Вторичная структура триозофосфатизомеразы и домена пируваткиназы
Рис. 1.6. Вторичная структура триозофосфатизомеразы и домена пируваткиназы
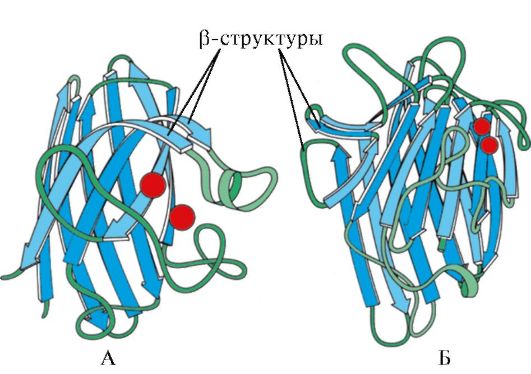 Рис. 1.7. Вторичная структура константного домена иммуноглобулина (А) и фермента супероксиддисмутазы (Б)
Рис. 1.7. Вторичная структура константного домена иммуноглобулина (А) и фермента супероксиддисмутазы (Б)
В четвертую категорию включены белки, имеющие в своем составе незначительное количество регулярных вторичных структур. К таким белкам можно отнести небольшие, богатые цистеином белки или металлопротеины.
Третичная структура белка - тип конформации, образующийся за счет взаимодействий между радикалами аминокислот, которые могут находиться на значительном расстоянии друг от друга в пептидной цепи. Большинство белков при этом формируют пространственную структуру, напоминающую глобулу (глобулярные белки).
Так как гидрофобные радикалы аминокислот имеют тенденцию к объединению с помощью так называемых гидрофобных взаимодействий и межмолекулярных ван-дер-ваальсовых сил, внутри белковой глобулы образуется плотное гидрофобное ядро. Гидрофильные ионизированные и неионизированные радикалы в основном располагаются на поверхности белка и определяют его растворимость в воде.
 Рис. 1.8. Типы связей, возникающих между радикалами аминокислот при формировании третичной структуры белка
Рис. 1.8. Типы связей, возникающих между радикалами аминокислот при формировании третичной структуры белка
1 - ионная связь - возникает между положительно и отрицательно заряженными функциональными группами;
2 - водородная связь - возникает между гидрофильной незаряженной и любой другой гидрофильной группой;
3 - гидрофобные взаимодействия - возникают между гидрофобными радикалами;
4 - дисульфидная связь - формируется за счет окисления SH-групп остатков цистеина и их взаимодействия друг с другом
Гидрофильные аминокислотные остатки, оказавшиеся внутри гидрофобного ядра, могут взаимодействовать друг с другом с помощью ионных и водородных связей (рис. 1.8).
Ионные и водородные связи, а также гидрофобные взаимодействия относятся к числу слабых: их энергия ненамного превышает энергию теплового движения молекул при комнатной температуре. Конформация белка поддерживается за счет возникновения множества таких слабых связей. Так как атомы, из которых состоит белок, находятся в постоянном движении, то возможен разрыв одних слабых связей и образование других, что приводит к небольшим перемещениям отдельных участков полипептидной цепи. Это свойство белков изменять конформацию в результате разрыва одних и образования других слабых связей называется конформационной лабильностью.
В организме человека функционируют системы, поддерживающие гомеостаз - постоянство внутренней среды в определенных допустимых для здорового организма пределах. В условиях гомеостаза небольшие изменения конформации не нарушают общую структуру и функцию белков. Функционально активная конформация белка называется нативной конформацией. Изменение внутренней среды (например, концентрации глюкозы, ионов Са, протонов и т.д.) приводит к изменению конформации и нарушению функций белков.
Третичная структура некоторых белков стабилизирована дисульфидными связями, образующимися за счет взаимодействия -SH групп двух остатков
 Рис. 1.9. Образование дисульфидной связи в молекуле белка
Рис. 1.9. Образование дисульфидной связи в молекуле белка
цистеина (рис. 1.9). Большинство внутриклеточных белков не имеет в третичной структуре ковалентных дисульфидных связей. Их наличие характерно для секретируемых клеткой белков, что обеспечивает их большую стабильность во внеклеточных условиях. Так, дисульфидные связи имеются в молекулах инсулина и иммуноглобулинов.
Инсулин - белковый гормон, синтезирующийся в β-клетках поджелудочной железы и секретируемый в кровь в ответ на повышение концентрации глюкозы в крови. В структуре инсулина имеются две дисульфидные связи, соединяющие полипептидные А- и В-цепи, и одна дисульфидная связь внутри А-цепи (рис. 1.10).
 Рис. 1.10. Дисульфидные связи в структуре инсулина
Рис. 1.10. Дисульфидные связи в структуре инсулина
5. Супервторичная структура белков. В разных по первичной структуре и функциям белках иногда выявляются сходные сочетания и взаиморасположение вторичных структур, которые называются супервторичной структурой. Она занимает промежуточное положение между вторичной и третичной структурами, поскольку это специфическое сочетание элементов вторичной структуры при формировании третичной структуры белка. Супервторичные структуры имеют специфические названия, такие как «α-спираль-поворот-а-спираль», «лейциновая застежка молния», «цинковые пальцы» и др. Такие супервторичные структуры характерны для ДНК-связывающих белков.
«Лейциновая застежка-молния». Этот вид супервторичной структуры используется для соединения двух белков. На поверхности взаимодействующих белков имеются α-спиральные участки, содержащие не менее четырех остатков лейцина. Лейциновые остатки в α-спирали располагаются через шесть аминокислот один от другого. Так как каждый виток α-спирали содержит 3,6 аминокислотных остатка, радикалы лейцина находятся на поверхности каждого второго витка. Лейциновые остатки α-спирали одного белка могут взаимодействовать с лейциновыми остатками другого белка (гидрофобные взаимодействия), соединяя их вместе (рис. 1.11.). Многие ДНК связывающие белки функционируют в составе олигомерных комплексов, где отдельные субъединицы связываются друг с другом «лейциновыми застежками».
 Рис. 1.11. «Лейциновая застежка-молния» между α-спиральными участками двух белков
Рис. 1.11. «Лейциновая застежка-молния» между α-спиральными участками двух белков
Примером таких белков могут служить гистоны. Гистоны - ядерные белки, в состав которых входит большое количество положительно заряженных аминокислот - аргинина и лизина (до 80%). Молекулы гистонов объединяются в олигомерные комплексы, содержащие восемь мономеров с помощью «лейциновых застежек», несмотря на значительный одноименный заряд этих молекул.
«Цинковый палец» - вариант супервторичной структуры, характерный для ДНК-связывающих белков, имеет вид вытянутого фрагмента на поверхности белка и содержит около 20 аминокислотных остатков (рис. 1.12). Форму «вытянутого пальца» поддерживает атом цинка, связанный с радикалами четыре аминокислот - двух остатков цистеина и двух - гистидина. В некоторых случаях вместо остатков гистидина находятся остатки цистеина. Два близко лежащих остатка цистеина отделены от двух других остатков Гисили Циспоследовательностью, состоящей примерно из 12 аминокислотных остатков. Этот участок белка образует α-спираль, радикалы которой могут специфично связываться с регуляторными участками большой бороздки ДНК. Специфичность связывания индивидуального
 Рис.
1.12. Первичная структура участка ДНК-связывающих белков, формирующих
структуру «цинкового пальца» (буквами обозначены аминокислоты, входящие в
состав этой структуры)
Рис.
1.12. Первичная структура участка ДНК-связывающих белков, формирующих
структуру «цинкового пальца» (буквами обозначены аминокислоты, входящие в
состав этой структуры)
регуляторного ДНК-связывающего белка зависит от последовательности аминокислотных остатков, расположенных в области «цинкового пальца». Такие структуры содержат, в частности, рецепторы стероидных гормонов, участвующих в регуляции транскрипции (считывание информации с ДНК на РНК).
ТЕМА 1.2. ОСНОВЫ ФУНКЦИОНИРОВАНИЯ БЕЛКОВ. ЛЕКАРСТВА КАК ЛИГАНДЫ, ВЛИЯЮЩИЕ НА ФУНКЦИЮ БЕЛКОВ
1. Активный центр белка и его взаимодействие с лигандом. В процессе формирования третичной структуры на поверхности функционально активного белка, обычно в углублении, образуется участок, сформированный радикалами аминокислот, далеко стоящими друг от друга в первичной структуре. Этот участок, имеющий уникальное строение для данного белка и способный специфично взаимодействовать с определенной молекулой или группой похожих молекул, называется центром связывания белка с лигандом или активным центром. Лигандами называются молекулы, взаимодействующие с белками.
Высокая специфичность взаимодействия белка с лигандом обеспечивается комплементарностью структуры активного центра структуре лиганда.
Комплементарность - это пространственное и химическое соответствие взаимодействующих поверхностей. Активный центр должен не только пространственно соответствовать входящему в него лиганду, но и между функциональными группами радикалов, входящих в активный центр, и лигандом должны образоваться связи (ионные, водородные, а также гидрофобные взаимодействия), которые удерживают лиганд в активном центре (рис. 1.13).
 Рис. 1.13. Комплементарное взаимодействие белка с лигандом
Рис. 1.13. Комплементарное взаимодействие белка с лигандом
Некоторые лиганды, присоединяясь к активному центру белка, выполняют вспомогательную роль в функционировании белков. Такие лиганды называются кофакторами, а белки, имеющие в своем составе небелковую часть, - сложными белками (в отличие от простых белков, состоящих только из белковой части). Небелковая часть, прочно соединенная с белком, носит название простетической группы. Например, в составе миоглобина, гемоглобина и цитохромов содержится прочно прикрепленная к активному центру простетическая группа - гем, содержащий ион железа. Сложные белки, содержащие гем, называются гемопротеинами.
При присоединении к белкам специфических лигандов проявляется функция этих белков. Так, альбумин - важнейший белок плазмы крови - проявляет свою транспортную функцию, присоединяя к активному центру гидрофобные лиганды, такие как жирные кислоты, билирубин, некоторые лекарства и др. (рис. 1.14)
Лигандами, взаимодействующими с трехмерной структурой пептидной цепи, могут быть не только низкомолекулярные органические и неорганические молекулы, но и макромолекулы:
ДНК (рассмотренные выше примеры с ДНК-связывающими белками);
Полисахариды;
 Рис. 1.14. Взаимосвязь генотипа и фенотипа
Рис. 1.14. Взаимосвязь генотипа и фенотипа
Уникальная первичная структура белков человека, закодированная в молекуле ДНК, в клетках реализуется в виде уникальной конформации, структуры активного центра и функций белков
В этих случаях белок узнает определенный участок лиганда, соразмерный и комплементарный центру связывания. Так на поверхности гепатоцитов имеются белки-рецепторы к гормону инсулину, имеющему также белковое строение. Взаимодействие инсулина с рецептором вызывает изменение его конформации и активации сигнальных систем, приводящих к запасанию в гепатоцитах питательных веществ после еды.
Таким образом, в основе функционирования белков лежит специфическое взаимодействие активного центра белка с лигандом.
2. Доменная структура и ее роль в функционировании белков. Длинные полипептидные цепи глобулярных белков часто складываются в несколько компактных, относительно независимых областей. Они имеют самостоятельную третичную структуру, напоминающую таковую у глобулярных белков, и называются доменами. Благодаря доменной структуре белков легче формируется их третичная структура.
В доменных белках центры связывания с лигандом часто располагаются между доменами. Так, трипсин - протеолитический фермент, который вырабатывается экзокринной частью поджелудочной железы и необходим для переваривания белков пищи. Он имеет двухдоменное строение, а центр связывания трипсина с его лигандом - пищевым белком - располагается в бороздке между двумя доменами. В активном центре создаются условия, необходимые для эффективного связывания специфического участка пищевого белка и гидролиза его пептидных связей.
Разные домены в белке при взаимодействии активного центра с лигандом могут перемещаться друг относительно друга (рис. 1.15).
Гексокиназа - фермент, катализирующий фосфорилирование глюкозы с помощью АТФ. Активный центр фермента располагается в расщелине между двумя доменами. При связывании гексокиназы с глюкозой окружающие ее домены смыкаются и субстрат оказывается в «ловушке», где и происходит фосфорилирование (см. рис. 1.15).
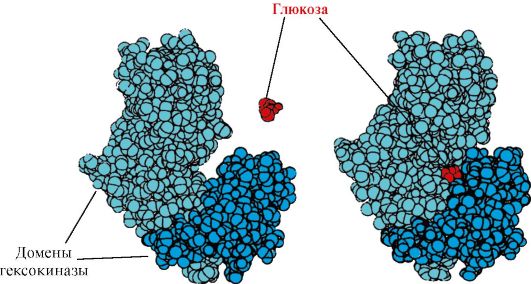 Рис. 1.15. Связывание доменов гексокиназы с глюкозой
Рис. 1.15. Связывание доменов гексокиназы с глюкозой
В некоторых белках домены выполняют самостоятельные функции, связываясь с различными лигандами. Такие белки называются многофункциональными.
3. Лекарства - лиганды, влияющие на функцию белков. Взаимодействие белков с лигандами специфично. Однако благодаря конформационной лабильности белка и его активного центра можно подобрать другое вещество, которое также могло бы взаимодействовать с белком в активном центре или ином участке молекулы.
Вещество, по структуре похожее на природный лиганд, называют структурным аналогом лиганда или неприродным лигандом. Оно также взаимодействует с белком в активном центре. Структурный аналог лиганда может как усиливать функцию белка (агонист), так и снижать ее (антагонист). Лиганд и его структурные аналоги конкурируют друг с другом за связывание с белком в одном центре. Такие вещества называются конкурентными модуляторами (регуляторами) белковых функций. Многие лекарственные препараты действуют как ингибиторы белков. Некоторые из них получают химической модификацией природных лигандов. Ингибиторы белковых функций могут быть лекарствами и ядами.
Атропин - конкурентный ингибитор М-холинорецепторов. Ацетилхолин - нейромедиатор передачи нервного импульса через холинэргические синапсы. Для проведения возбуждения выделившийся в синаптическую щель ацетилхолин должен взаимодействовать с белком - рецептором постсинаптической мембраны. Обнаружены два типа холинорецепторов:
М-рецептор, кроме ацетилхолина избирательно взаимодействующий с мускарином (токсином мухомора). М - холинорецепторы имеются на гладких мышцах и при взаимодействии с ацетилхолином вызывают их сокращение;
Н-рецептор, специфично связывающийся с никотином. Н-холинорецепторы обнаружены в синапсах поперечнополосатых скелетных мышц.
Специфическим ингибитором М-холинорецепторов является атропин. Он содержится в растениях красавке и белене.
 Атропин
имеет в структуре схожие с ацетилхолином функциональные группы и их
пространственное расположение, поэтому относится к конкурентным
ингибиторам М-холинорецепторов. Учитывая, что связывание ацетилхолина с
М-холинорецепторами вызывает сокращение гладких мышц, атропин используют
как лекарство, снимающее их спазм (спазмолитик).
Так, известно
применение атропина для расслабления глазных мышц при просмотре глазного
дна, а также для снятия спазмов при желудочнокишечных коликах.
М-холинорецепторы имеются и в центральной нервной системе (ЦНС), поэтому
большие дозы атропина могут вызвать нежелательную реакцию со стороны
ЦНС: двигательное и психическое возбуждение, галлюцинации, судороги.
Атропин
имеет в структуре схожие с ацетилхолином функциональные группы и их
пространственное расположение, поэтому относится к конкурентным
ингибиторам М-холинорецепторов. Учитывая, что связывание ацетилхолина с
М-холинорецепторами вызывает сокращение гладких мышц, атропин используют
как лекарство, снимающее их спазм (спазмолитик).
Так, известно
применение атропина для расслабления глазных мышц при просмотре глазного
дна, а также для снятия спазмов при желудочнокишечных коликах.
М-холинорецепторы имеются и в центральной нервной системе (ЦНС), поэтому
большие дозы атропина могут вызвать нежелательную реакцию со стороны
ЦНС: двигательное и психическое возбуждение, галлюцинации, судороги.
Дитилин - конкурентный агонист Н-холинорецепторов, ингибирующий функцию нервно-мышечных синапсов.
 Нервно-мышечные
синапсы скелетных мышц содержат Н-холинорецепторы. Их взаимодействие с
ацетилхолином приводит к мышечным сокращениям. При некоторых
хирургических операциях, а также в эндоскопических исследованиях
используют препараты, вызывающие расслабление скелетных мышц (миорелаксанты).
К
ним относится дитилин, являющийся структурным аналогом ацетилхолина. Он
присоединяется к Н-холинорецепторам, но в отличие от ацетилхолина очень
медленно разрушается ферментом - ацетилхолинэстеразой. В результате
длительного открытия ионных каналов и стойкой деполяризации мембраны
нарушается проведение нервного импульса и происходит мышечное
расслабление. Первоначально эти свойства были обнаружены у яда кураре,
поэтому такие препараты называют курареподобными.
Нервно-мышечные
синапсы скелетных мышц содержат Н-холинорецепторы. Их взаимодействие с
ацетилхолином приводит к мышечным сокращениям. При некоторых
хирургических операциях, а также в эндоскопических исследованиях
используют препараты, вызывающие расслабление скелетных мышц (миорелаксанты).
К
ним относится дитилин, являющийся структурным аналогом ацетилхолина. Он
присоединяется к Н-холинорецепторам, но в отличие от ацетилхолина очень
медленно разрушается ферментом - ацетилхолинэстеразой. В результате
длительного открытия ионных каналов и стойкой деполяризации мембраны
нарушается проведение нервного импульса и происходит мышечное
расслабление. Первоначально эти свойства были обнаружены у яда кураре,
поэтому такие препараты называют курареподобными.
ТЕМА 1.3. ДЕНАТУРАЦИЯ БЕЛКОВ И ВОЗМОЖНОСТЬ ИХ СПОНТАННОЙ РЕНАТИВАЦИИ
1. Так как нативная конформация белков поддерживается за счет слабых взаимодействий, изменение состава и свойств окружающей белок среды, воздействие химических реагентов и физических факторов вызывают изменение их конформации (свойство конформационной лабильности). Разрыв большого количества связей приводит к разрушению нативной конформации и денатурации белков.
Денатурация белков - это разрушение их нативной конформации под действием денатурирующих агентов, вызванное разрывом слабых связей, стабилизирующих пространственную структуру белка. Денатурация сопровождается разрушением уникальной трехмерной структуры и активного центра белка и потерей его биологической активности (рис. 1.16).
Все денатурированные молекулы одного белка приобретают случайную конформацию, отличающуюся от других молекул того же белка. Радикалы аминокислот, формирующие активный центр, оказываются пространственно удаленными друг от друга, т.е. разрушается специфический центр связывания белка с лигандом. При денатурации первичная структура белков остается неизменной.
Применение денатурирующих агентов в биологических исследованиях и медицине. В биохимических исследованиях перед определением в биологическом материале низкомолекулярных соединений обычно из раствора вначале удаляют белки. Для этой цели чаще всего используют трихлоруксусную кислоту (ТХУ). После добавления ТХУ в раствор денатурированные белки выпадают в осадок и легко удаляются фильтрованием (табл. 1.1.)
В медицине денатурирующие агенты часто применяют для стерилизации медицинского инструмента и материала в автоклавах (денатурирующий агент - высокая температура) и в качестве антисептиков (спирт, фенол, хлорамин) для обработки загрязненных поверхностей, содержащих патогенную микрофлору.
2. Спонтанная ренативация белков - доказательство детерминированности первичной структуры, конформации и функции белков. Индивидуальные белки - это продукты одного гена, которые имеют идентичную аминокислотную последовательность и в клетке приобретают одинаковую конформацию. Фундаментальный вывод о том, что в первичной структуре белка уже заложена информация о его конформации и функции, был сделан на основе способности некоторых белков (в частности, рибонуклеазы и миоглобина) к спонтанной ренативации - восстановлению их нативной конформации после денатурации.
Формирование пространственных структур белка осуществляется способом самосборки - самопроизвольного процесса, при котором полипептидная цепь, имеющая уникальную первичную структуру, стремится принять в растворе конформацию с наименьшей свободной энергией. Способность к ренативации белков, сохраняющих после денатурации первичную структуру, описана в опыте с ферментом рибонуклеазой.
Рибонуклеаза - фермент, разрушающий связи между отдельными нуклеотидами в молекуле РНК. Этот глобулярный белок имеет одну полипептидную цепь, третичная структура которой стабилизирована множеством слабых и четырьмя дисульфидными связями.
Обработка рибонуклеазы мочевиной, разрушающей водородные связи в молекуле, и восстановителем, разрывающим дисульфидные связи, приводит к денатурации фермента и потере его активности.
Удаление денатурирующих агентов диализом приводит к восстановлению конформации и функции белка, т.е. к ренативации. (рис. 1.17).
 Рис. 1.17. Денатурация и ренативация рибонуклеазы
Рис. 1.17. Денатурация и ренативация рибонуклеазы
А - нативная конформация рибонуклеазы, в третичной структуре которой имеются четыре дисульфидные связи; Б - денатурированная молекула рибонуклеазы;
В - ренативированная молекула рибонуклеазы с восстановленной структурой и функцией
1. Заполните таблицу 1.2.
Таблица 1.2. Классификация аминокислот по полярности радикалов
2. Напишите формулу тетрапептида:
Асп - Про - Фен - Лиз
а) выделите в пептиде повторяющиеся группы, образующие пептидный остов, и вариабельные группы, представленные радикалами аминокислот;
б) обозначьте N- и С-концы;
в) подчеркните пептидные связи;
г) напишите другой пептид, состоящий из тех же аминокислот;
д) подсчитайте количество возможных вариантов тетрапептида с аналогичным аминокислотным составом.
3. Объясните роль первичной структуры белков на примере сравнительного анализа двух сходных по структуре и эволюционно близких пептидных гормонов нейрогипофиза млекопитающих - окситоцина и вазопрессина (табл. 1.3).
Таблица 1.3. Структура и функции окситоцина и вазопрессина
 Для этого:
Для этого:
а) сравните состав и последовательность аминокислот двух пептидов;
б) найдите сходство первичной структуры двух пептидов и сходство их биологического действия;
в) найдите различия в структуре двух пептидов и различие их функций;
г) сделайте вывод о влиянии первичной структуры пептидов на их функции.
4. Опишите основные этапы формирования конформации глобулярных белков (вторичная, третичная структуры, понятие о супервторичной структуре). Укажите типы связей, участвующих в формировании структур белка. Радикалы каких аминокислот могут участвовать в образовании гидрофобных взаимодействий, ионных, водородных связях.
Приведите примеры.
5. Дайте определение понятию «конформационная лабильность белков», укажите причины ее существования и значение.
6. Раскройте смысл следующей фразы: «В основе функционирования белков лежит их специфическое взаимодействие с лигандом», используя термины и объясняя их значение: конформация белка, активный центр, лиганд, комплементарность, функция белка.
7. На одном из примеров объясните, что такое домены и какова их роль в функционировании белков.
ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ
1. Установите соответствие.
Функциональная группа в радикале аминокислоты:
А. Карбоксильная группа Б. Гидроксильная группа В Гуанидиновая группа Г. Тиольная группа Д. Аминогруппа
2. Выберите правильные ответы.
Аминокислоты с полярными незаряженными радикалами - это:
A. Цис Б. Асн
B. Глу Г. Три
3. Выберите правильные ответы.
Радикалы аминокислот:
A. Обеспечивают специфичность первичной структуры Б. Участвуют в формировании третичной структуры
B. Располагаясь на поверхности белка, влияют на его растворимость Г. Формируют активный центр
Д. Участвуют в образовании пептидных связей
4. Выберите правильные ответы.
Гидрофобные взаимодействия могут образовываться между радикалами аминокислот:
A. Тре Лей Б. Про Три
B. Мет Иле Г. Тир Ала Д. Вал Фен
5. Выберите правильные ответы.
Ионные связи могут образовываться между радикалами аминокислот:
A. Глн Асп Б. Apr Лиз
B. Лиз Глу Г. Гис Асп Д. Асн Apr
6. Выберите правильные ответы.
Водородные связи могут образовываться между радикалами аминокислот:
A. Сер Глн Б. Цис Тре
B. Асп Лиз Г. Глу Асп Д. Асн Тре
7. Установите соответствие.
Тип связи, участвующий в формировании структуры белка:
A. Первичная структура Б. Вторичная структура
B. Третичная структура
Г. Супервторичная структура Д. Конформация.
1. Водородные связи между атомами пептидного остова
2. Слабые связи между функциональными группами радикалов аминокислот
3. Связи между α-амино и α-карбоксильными группами аминокислот
8. Выберите правильные ответы. Трипсин:
A. Протеолитический фермент Б. Содержит два домена
B. Гидролизирует крахмал
Г. Активный центр расположен между доменами. Д. Состоит из двух полипептидных цепей.
9. Выберите правильные ответы. Атропин:
A. Нейромедиатор
Б. Структурный аналог ацетилхолина
B. Взаимодействует с Н-холинорецепторами
Г. Усиливает проведение нервного импульса через холинэргические синапсы
Д. Конкурентный ингибитор М-холинорецепторов
10. Выберите правильные утверждения. В белках:
A. Первичная структура содержит информацию о строении его активного центра
Б. Активный центр формируется на уровне первичной структуры
B. Конформация жестко фиксирована ковалентными связями
Г. Активный центр может взаимодействовать с группой похожих лигандов
благодаря конформационной лабильности белков Д. Изменение окружающей среды, может влиять на сродство активного
центра к лиганду
1. 1-В, 2-Г, 3-Б.
3. А, Б, В, Г.
7. 1-Б, 2-Д, 3-А.
8. А, Б, В, Г.
ОСНОВНЫЕ ТЕРМИНЫ И ПОНЯТИЯ
1. Белок, полипептид, аминокислоты
2. Первичная, вторичная, третичная структуры белка
3. Конформация, нативная конформация белка
4. Ковалентные и слабые связи в белке
5. Конформационная лабильность
6. Активный центр белка
7. Лиганды
8. Фолдинг белков
9. Структурные аналоги лигандов
10. Доменные белки
11. Простые и сложные белки
12. Денатурация белка, денатурирующие агенты
13. Ренативация белков
Решите задачи
«Структурная организация белков и основы их функционирования»
1. Основная функция белка - гемоглобина А (НвА) - транспорт кислорода к тканям. В популяции людей известны множественные формы этого белка с измененными свойствами и функцией - так называемые аномальные гемоглобины. Например, установлено, что гемоглобин S, обнаруженный в эритроцитах больных серповидно-клеточной анемией (HbS), имеет низкую растворимость в условиях низкого парциального давления кислорода (как это имеет место в венозной крови). Это приводит к образованию агрегатов данного белка. Белок утрачивает свою функцию, выпадает в осадок, а эритроциты приобретают неправильную форму (некоторые из них образуют форму серпа) и быстрее обычного разрушаются в селезенке. В результате развивается серповидноклеточная анемия.
Единственное различие в первичной структуре НвА и обнаружено в N-концевом участке β-цепи гемоглобина. Сравните N-концевые участки β-цепи и покажите, как изменения в первичной структуре белка влияют на его свойства и функции.
 Для этого:
Для этого:
а) напишите формулы аминокислот, по которым различаются НвА и сравните свойства этих аминокислот (полярность, заряд).
б) сделайте вывод о причине снижения растворимости и нарушении транспорта кислорода в ткани.
2. На рисунке представлена схема строения белка, имеющего центр связывания с лигандом (активный центр). Объясните, почему белок обладает избирательностью в выборе лиганда. Для этого:
а) вспомните, что такое активный центр белка, и рассмотрите строение активного центра белка, представленного на рисунке;
б) напишите формулы радикалов аминокислот, входящих в состав активного центра;
в) нарисуйте лиганд, который мог бы специфически взаимодействовать с активным центром белка. Укажите на нем функциональные группы, способные образовать связи с радикалами аминокислот, входящих в состав активного центра;
г) укажите типы связей, возникающих между лигандом и радикалами аминокислот активного центра;
д) объясните, на чем основана специфичность взаимодействия белка с лигандом.
 3.
На рисунке представлен активный центр белка и несколько лигандов.
3.
На рисунке представлен активный центр белка и несколько лигандов.
Определите, какой из лигандов с наибольшей вероятностью будет взаимодействовать с активным центром белка и почему.
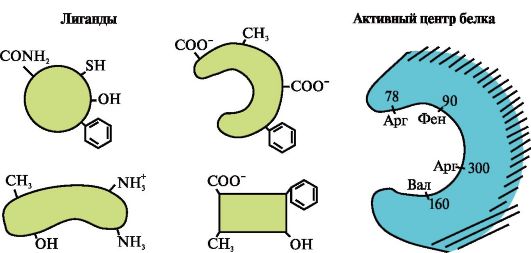 Какие типы связей возникают в процессе образования комплекса белок-лиганд»?
Какие типы связей возникают в процессе образования комплекса белок-лиганд»?
4. Структурные аналоги естественных лигандов белков могут использоваться в качестве лекарственных препаратов для изменения активности белков.
Ацетилхолин - медиатор передачи возбуждения в нервно-мышечных синапсах. При взаимодействии ацетилхолина с белками - рецепторами постсинаптической мембраны скелетных мышц происходит открытие ионных каналов и мышечное сокращение. Дитилин - лекарство, применяемое при некоторых операциях для расслабления мышц, так как он нарушает передачу нервного импульса через нервно-мышечные синапсы. Объясните механизм действия дитилина как миорелаксирующего препарата. Для этого:
а) напишите формулы ацетилхолина и дитилина и сравните их структуры;
б) опишите механизм расслабляющего действия дитилина.
5. При некоторых заболеваниях у больного повышается температура тела, что рассматривают как защитную реакцию организма. Однако высокие температуры губительны для белков организма. Объясните, почему при температуре выше 40 °С нарушается функция белков и возникает угроза для жизни человека. Для этого вспомните:
1) Строение белков и связи, удерживающие его структуру в нативной конформации;
2) Как меняется структура и функция белков при повышении температуры?;
3) Что такое гомеостаз и почему он важен для поддержания здоровья человека.
Модульная единица 2 ОЛИГОМЕРНЫЕ БЕЛКИ КАК МИШЕНИ РЕГУЛЯТОРНЫХ ВОЗДЕЙСТВИЙ. СТРУКТУРНО-ФУНКЦИОНАЛЬНОЕ МНОГООБРАЗИЕ БЕЛКОВ. МЕТОДЫ РАЗДЕЛЕНИЯ И ОЧИСТКИ БЕЛКОВ
Цели изучения Уметь:
1. Использовать знания об особенностях структуры и функций олигомерных белков для понимания адаптивных механизмов регуляции их функций.
2. Объяснять роль шаперонов в синтезе и поддержании конформации белков в условиях клетки.
3. Объяснять многообразие проявления жизни многообразием структур и функций синтезирующихся в организме белков.
4. Анализировать связь структуры белков с их функцией на примерах сравнения родственных гемопротеинов - миоглобина и гемоглобина, а также представителей пяти классов белков семейства иммуноглобулинов.
5. Применять знания об особенностях физико-химических свойств белков для выбора методов их очистки от других белков и примесей.
6. Интерпретировать результаты количественного и качественного состава белков плазмы крови для подтверждения или уточнения клинического диагноза.
Знать:
1. Особенности строения олигомерных белков и адаптивные механизмы регуляции их функций на примере гемоглобина.
2. Строение и функции шаперонов и их значение для поддержания нативной конформации белков в условиях клетки.
3. Принципы объединения белков в семейства по схожести их конформации и функций на примере иммуноглобулинов.
4. Методы разделения белков, основанные на особенностях их физикохимических свойств.
5. Электрофорез плазмы крови как метод оценки качественного и количественного состава белков.
ТЕМА 1.4. ОСОБЕННОСТИ СТРОЕНИЯ И ФУНКЦИОНИРОВАНИЯ ОЛИГОМЕРНЫХ БЕЛКОВ НА ПРИМЕРЕ ГЕМОГЛОБИНА
1. Многие белки имеют в своем составе несколько полипептидных цепей. Такие белки называют олигомерными, а отдельные цепи - протомерами. Протомеры в олигомерных белках соединены множеством слабых нековалентных связей (гидрофобных, ионных, водородных). Взаимодействие
протомеров осуществляется благодаря комплементарности их контактирующих поверхностей.
Количество протомеров в олигомерных белках может сильно варьировать: гемоглобин содержит 4 протомера, фермент аспартатаминотрансфераза - 12 протомеров, а в белок вируса табачной мозаики входит 2120 протомеров, соединенных нековалентными связями. Следовательно, олигомерные белки могут иметь очень большую молекулярную массу.
Взаимодействие одного протомера с другими можно рассматривать как частный случай взаимодействия белка с лигандом, так как каждый протомер служит лигандом для других протомеров. Количество и способ соединения протомеров в белке называется четвертичной структурой белка.
В состав белков могут входить одинаковые или разные по строению протомеры, например, гомодимеры - белки, содержащие два одинаковых протомера, а гетеродимеры - белки, содержащие два разных протомера.
Если в состав белков входят разные протомеры, то на них могут формироваться отличающиеся по структуре центры связывания с разными лигандами. При связывании лиганда с активным центром проявляется функция данного белка. Центр, расположенный на другом протомере, называется аллостерическим (другим, отличным от активного). Связываясь с аллостерическим лигандом или эффектором, он выполняет регуляторную функцию (рис. 1.18). Взаимодействие аллостерического центра с эффектором вызывает конформационные изменения в структуре всего олигомерного белка благодаря его конформационной лабильности. Это влияет на сродство активного центра к специфическому лиганду и регулирует функцию данного белка. Изменение конформации и функции всех протомеров при взаимодействии олигомерного белка хотя бы с одним лигандом носит название кооперативных изменений конформации. Эффекторы, усиливающие функцию белка, называются активаторами, а эффекторы, угнетающие его функцию, - ингибиторами.
Таким образом, у олигомерных белков, а также белков, имеющих доменное строение, появляется новое по сравнению с мономерными белками свойство - способность к аллостерической регуляции функций (регуляции присоединением к белку разных лигандов). Это можно проследить, сравнивая структуры и функции двух близко родственных сложных белков миоглобина и гемоглобина.
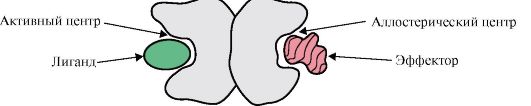 Рис. 1.18. Схема строения димерного белка
Рис. 1.18. Схема строения димерного белка
2. Формирование пространственных структур и функционирование миоглобина.
Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О 2 , необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe+ 2 гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О 2 , поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O 2 резко снижается.
Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых латинскими буквами от А до Н, между которыми имеются неспирализованные участки. Третичная структура Мв имеет вид компактной глобулы, в углублении которой между F и Е α-спиралями расположен активный центр (рис. 1.19).
 Рис. 1.19. Структура миоглобина
Рис. 1.19. Структура миоглобина
3. Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три 3 9 и Фен 138) К активному центру присоединяется плохо растворимые в воде лиганды - гем и О 2 . Гем - специфический лиганд апоМв (рис. 1.20), основу которого составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe+ 2 , соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е 7 (Гис 64) и Гис F 8 (Гис 93) (рис. 1.21).
 Рис. 1.20. Строение гема - небелковой части миоглобина и гемоглобина
Рис. 1.20. Строение гема - небелковой части миоглобина и гемоглобина
 Рис. 1.21. Расположение гема и O 2 в активном центре апомиоглобина и протомеров гемоглобина
Рис. 1.21. Расположение гема и O 2 в активном центре апомиоглобина и протомеров гемоглобина
Гем через атом железа ковалентно связан с Гис F 8 . O 2 присоединяется к железу с другой стороны плоскости гема. Гис Е 7 необходим для правильной ориентации О 2 и облегчает присоединение кислорода к Fe+ 2 гема
Гис F 8 образует координационную связь с Fe+ 2 и прочно фиксирует гем в активном центре. Гис Е 7 необходим для правильной ориентации в активном центре другого лиганда - O 2 при его взаимодействии с Fe+ 2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O 2 с Fe +2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе+ 3 .
Мономерное строение Мв и его активного центра определяет высокое сродство белка к О 2 .
4. Олигомерное строение Нв и регуляция сродства Нв к О 2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2р-цепи).
В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А.
Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями (рис 1.22).
Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F 8 и Гис Е 7 , которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом (см. строение миоглобина).
 Рис. 1.22. Олигомерная структура гемоглобина
Рис. 1.22. Олигомерная структура гемоглобина
Кроме того, Гис Е 7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О 2 . СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О 2 . Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О 2 всего в 200 раз благодаря наличию в активном центре Гис Е 7 . Остаток этой аминокислоты создает оптимальные условия для связывания гема с O 2 и ослабляет взаимодействие гема с СО.
5. Основная функция Нв - транспорт О 2 из легких в ткани. В отличие от мономерного миоглобина, имеющего очень высокое сродство к О 2 и выполняющего функцию запасания кислорода в красных мышцах, олигомерная структура гемоглобина обеспечивает:
1) быстрое насыщение Нв кислородом в легких;
2) способность Нв отдавать кислород в тканях при относительно высоком парциальном давлении O 2 (20-40 мм рт. ст.);
3) возможность регуляции сродства Нв к О 2 .
6. Кооперативные изменения конформации протомеров гемоглобина ускоряют связывание O 2 в легких и отдачу его в ткани. В легких высокое парциальное давление O 2 способствует связыванию его с Нв в активном центре четырех протомеров (2α и 2β). Активный центр каждого протомера, так же как и в миоглобине, расположен между двумя α-спиралями (F и Е) в гидрофобном кармане. Он содержит небелковую часть - гем, прикрепленный к белковой части множеством слабых гидрофобных взаимодействий и одной прочной связью между Fe 2 + гема и Гис F 8 (см. рис. 1.21).
В дезоксигемоглобине, благодаря этой связи с Гис F 8 , атом Fe 2 + выступает из плоскости гема по направлению к гистидину. Связывание O 2 с Fe 2 + происходит по другую сторону гема в области Гис Е 7 с помощью единственной свободной координационной связи. Гис Е 7 обеспечивает оптимальные условия для связывания O 2 с железом гема.
Присоединение O 2 к атому Fe +2 одного протомера вызывает его перемещение в плоскость гема, а за ним и остатка гистидина, связанного с ним
 Рис. 1.23. Изменение конформации протомера гемоглобина при соединении с O 2
Рис. 1.23. Изменение конформации протомера гемоглобина при соединении с O 2
Это приводит к изменению конформации всех полипептидных цепей за счет их конформационной лабильности. Изменение конформации других цепей облегчает их взаимодействие со следующими молекулами О 2 .
Четвертая молекула О 2 присоединяется к гемоглобину в 300 раз легче, чем первая (рис. 1.24).
 Рис. 1.24. Кооперативные изменения конформации протомеров гемоглобина при его взаимодействии с О 2
Рис. 1.24. Кооперативные изменения конформации протомеров гемоглобина при его взаимодействии с О 2
В тканях каждая следующая молекула O 2 отщепляется легче, чем предыдущая, также за счет кооперативных изменений конформации протомеров.
7. CO 2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О 2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O 2 , доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО 2 и K 2 O, количество которых пропорционально интенсивности протекающих процессов окисления.
СO 2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион.
Н+ способны присоединятся к радикалам Гис 14 6 в α- и β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О 2 , способствует отщеплению О 2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов (рис. 1.25).
Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект).
В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н + . Освободившиеся протоны под действием карбангидразы взаимодействуют с бикарбонатами с образованием СО 2 и Н 2 О

 Рис. 1.25. Зависимость сродства Нв к О 2 от концентрации СО 2 и протонов (эффект Бора):
Рис. 1.25. Зависимость сродства Нв к О 2 от концентрации СО 2 и протонов (эффект Бора):
А - влияние концентрации СО 2 и Н+ на высвобождение О 2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО 2 .
Образовавшийся СО 2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при физических нагрузках, тем выше концентрация СО 2 и Н + и тем больше кислорода получают ткани в результате уменьшения сродства Нв к О 2 .
8. Аллостерическая регуляция сродства Нв к О 2 лигандом - 2,3-бис- фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд -5.
 Бисфосфоглицерат
в капиллярах тканей, связываясь с дезоксигемоглобином, увеличивает
выход кислорода в ткани, уменьшая сродство Нв к О 2 .
Бисфосфоглицерат
в капиллярах тканей, связываясь с дезоксигемоглобином, увеличивает
выход кислорода в ткани, уменьшая сродство Нв к О 2 .
В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров (см. рис. 1.22). В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О 2 . В молекуле
дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. Из-за различия в размерах центральной полости 2,3-БФГ может присоединяться только к дезоксигемоглобину.
2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с пятью положительно заряженными группами двух β-цепей Нв: N-концевой α-аминогруппой Вал и радикалами Лиз 82 Гис 143 (рис. 1.26).
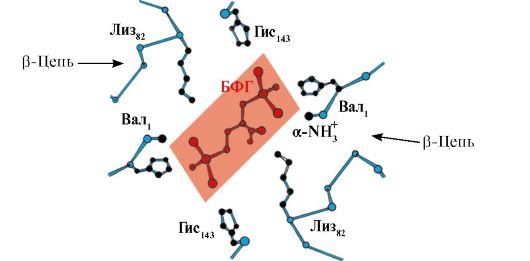 Рис. 1.26. БФГ в центральной полости дезоксигемоглобина
Рис. 1.26. БФГ в центральной полости дезоксигемоглобина
БФГ связывается с тремя положительно заряженными группами в каждой β-цепи.
В капиллярах тканей образующийся дезоксигемоглобин взаимодействует с 2,3-БФГ и между положительно заряженными радикалами β-цепей и отрицательно заряженным лигандом образуются ионные связи, которые изменяют конформацию белка и снижают сродство Нв к О 2 . Уменьшение сродства Нв к О 2 способствует более эффективному выходу О 2 в ткани.
В легких при высоком парциальном давлении кислород взаимодействует с Нв, присоединяясь к железу гема; при этом изменяется конформация белка, уменьшается центральная полость и происходит вытеснение 2,3-БФГ из аллостерического центра
Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках,
пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.
ТЕМА 1.5. ПОДДЕРЖАНИЕ НАТИВНОЙ КОНФОРМАЦИИ БЕЛКОВ В УСЛОВИЯХ КЛЕТКИ
В клетках в процессе синтеза полипептидных цепей, их транспорта через мембраны в соответствующие отделы клетки, в процессе фолдинга (формирования нативной конформации) и при сборке олигомерных белков, а также в период их функционирования в структуре белков возникают промежуточные, склонные к агрегации, нестабильные конформации. Гидрофобные радикалы, в нативной конформации обычно спрятанные внутри белковой молекулы, в нестабильной конформации оказываются на поверхности и стремятся к объединению с такими же плохо растворимыми в воде группами других белков. В клетках всех известных организмов обнаружены специальные белки, которые обеспечивают оптимальный фолдинг белков клетки, стабилизируют их нативную конформацию при функционировании и, что особенно важно, поддерживают структуру и функции внутриклеточных белков при нарушении гомеостаза. Эти белки получили название «шапероны», что в переводе с французского обозначает «няня».
1. Молекулярные шапероны и их роль в предотвращении денатурации белков.
Шапероны (Ш) классифицируются по массе субъединиц. Высокомолекулярные шапероны имеют массу от 60 до 110 кД. Среди них наиболее изучены три класса: Ш-60, Ш-70 и Ш-90. Каждый класс включает семейство родственных белков. Так, в состав Ш-70 входят белки с молекулярной массой от 66 до 78 кД. Низкомолекулярные шапероны имеют молекулярную массу от 40 до 15 кД.
Среди шаперонов различают конститутивные белки, высокий базальный синтез которых не зависит от стрессовых воздействий на клетки организма, и индуцибельные, синтез которых в нормальных условиях идет слабо, но резко возрастает при стрессовых воздействиях. Индуцибельные шапероны называют также «белками теплового шока», так как впервые они были обнаружены в клетках, подвергавшихся воздействию высоких температур. В клетках из-за высокой концентрации белков самопроизвольная ренативация частично денатурированных белков затруднена. Ш-70 могут предотвращать начавшийся процесс денатурации и способствовать восстановлению нативной конформации белков. Молекулярные шапероны-70 - высококонсервативный класс белков, находящихся во всех отделах клетки: цитоплазме, ядре, эндоплазматическом ретикулуме, митохондриях. На карбоксильном конце единственной полипептидной цепи Ш-70 имеется участок, который представляет собой бороздку, способный взаимодействовать с пептидами длиной
от 7 до 9 аминокислотных остатков, обогащенных гидрофобными радикалами. Такие участки в глобулярных белках встречаются примерно через каждые 16 аминокислот. Ш-70 способны защищать белки от температурной инактивации и восстанавливать конформацию и активность частично денатурированных белков.
2. Роль шаперонов в фолдинге белков. При синтезе белков на рибосоме N-концевая область полипептида синтезируется раньше С-концевой. Для формирования нативной конформации необходима полная аминокислотная последовательность белка. В процессе синтеза белков шапероны-70, благодаря строению их активного центра, способны закрывать склонные к агрегации участки полипептида, обогащенные гидрофобными радикалами аминокислот до завершения синтеза (рис 1.27, А).
 Рис. 1.27. Участие шаперонов в фолдинге белков
Рис. 1.27. Участие шаперонов в фолдинге белков
А - участие шаперонов-70 в предотвращении гидрофобных взаимодействий между участками синтезирующегося полипептида; Б - формирование нативной конформации белка в шапероновом комплексе
Многие высокомолекулярные белки, имеющие сложную конформацию, например доменное строение, осуществляют фолдинг в специальном пространстве, сформированном Ш-60. Ш-60 функционируют в виде олигомерного комплекса, состоящего из 14 субъединиц. Они формируют два полых кольца, каждое из которых состоит из семи субъединиц, эти кольца соединены друг с другом. Каждая субъединица Ш-60 состоит из трех доменов: апикального (верхушечного), обогащенного гидрофобными радикалами, обращенными в полость кольца, промежуточного и экваториального (рис. 1.28).
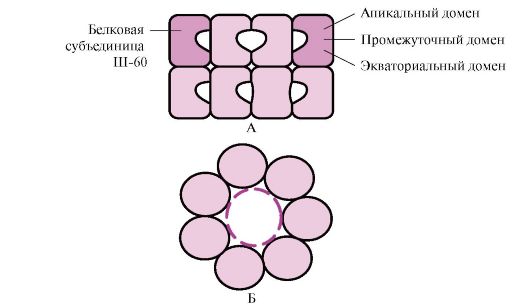 Рис. 1.28 . Структура шаперонинового комплекса, состоящего из 14 Ш-60
Рис. 1.28 . Структура шаперонинового комплекса, состоящего из 14 Ш-60
А - вид сбоку; Б - вид сверху
Синтезированные белки, имеющие на поверхности элементы, характерные для несвернутых молекул, в частности гидрофобные радикалы, попадают в полость шапероновых колец. В специфической среде этих полостей происходит перебор возможных конформаций, пока не будет найдена единственная, энергетически наиболее выгодная (рис. 1.27, Б). Формирование конформаций и высвобождение белка сопровождается гидролизом АТФ в экваториальной области. Обычно такой шаперонозависимый фолдинг требует затрат значительного количества энергии.
Кроме участия в формировании трехмерной структуры белков и ренативации частично денатурированных белков, шапероны также необходимы для протекания таких фундаментальных процессов, как сборка олигомерных белков, узнавание и транспорт в лизосомы денатурированных белков, транспорт белков через мембраны, участие в регуляции активности белковых комплексов.
ТЕМА 1.6. МНОГООБРАЗИЕ БЕЛКОВ. СЕМЕЙСТВА БЕЛКОВ НА ПРИМЕРЕ ИММУНОГЛОБУЛИНОВ
1. Белки играют решающую роль в жизнедеятельности отдельных клеток и всего многоклеточного организма, а их функции удивительно многообразны. Это определяется особенностями первичной структуры и конформаций белков, уникальностью строения активного центра и способностью связывать специфические лиганды.
Лишь очень небольшая часть всех возможных вариантов пептидных цепей может принять стабильную пространственную структуру; большинство
из них может принимать множество конформаций с примерно одинаковой энергией Гиббса, но с различными свойствами. Первичная структура большинства известных белков, отобранных биологической эволюцией, обеспечивает исключительную стабильность одной из конформаций, которая определяет особенности функционирования этого белка.
2. Семейства белков. В пределах одного биологического вида замены аминокислотных остатков могут приводить к возникновению разных белков, выполняющих родственные функции и имеющих гомологичные последовательности аминокислот. Такие родственные белки имеют поразительно сходные конформации: количество и взаиморасположение α-спиралей и (или) β-структур, большинство поворотов и изгибов полипептидных цепей похожи или идентичны. Белки с гомологичными участками полипептидной цепи, сходной конформацией и родственными функциями выделяют в семейства белков. Примеры семейств белков: сериновые протеиназы, семейство иммуноглобулинов, семейство миоглобина.
Сериновые протеиназы - семейство белков, выполняющих функцию протеолитических ферментов. К ним относятся пищеварительные ферменты - химотрипсин, трипсин, эластаза и многие факторы свертывания крови. Эти белки имеют в 40% положений идентичные аминокислоты и очень близкую конформацию (рис. 1.29).

Рис. 1.29 . Пространственные структуры эластазы (А) и химотрипсина (Б)
Некоторые аминокислотные замены привели к изменению субстратной специфичности этих белков и возникновению функционального многообразия внутри семейства.
3. Семейство иммуноглобулинов. В работе иммунной системы огромную роль играют белки суперсемейства иммуноглобулинов, которое включает в себя три семейства белков:
Антитела (иммуноглобулины);
Рецепторы Т-лимфоцитов;
Белки главного комплекса гистосовместимости - МНС 1-го и 2-го классов (Major Histocompatibility Complex).
Все эти белки имеют доменное строение, состоят из гомологичных иммуноподобных доменов и выполняют сходные функции: взаимодействуют с чужеродными структурами, либо растворенными в крови, лимфе или межклеточной жидкости (антитела), либо находящимися на поверхности клеток (собственных или чужеродных).
4. Антитела - специфические белки, вырабатываемые В-лимфоцитами в ответ на попадание в организм чужеродной структуры, называемой антигеном.
Особенности строения антител
Простейшие молекулы антител состоят из четырех полипептидных цепей: двух идентичных легких - L, содержащих около 220 аминокислот, и двух идентичных тяжелых - Н, состоящих из 440-700 аминокислот. Все четыре цепи в молекуле антитела соединены множеством нековалентных связей и четырьмя дисульфидными связями (рис. 1.30).
Легкие цепи антитела состоят из двух доменов: вариабельного (VL), находящегося в N-концевой области полипептидной цепи, и константного (CL), расположенного на С-конце. Тяжелые цепи обычно имеют четыре домена: один вариабельный (VH), находящийся на N-конце, и три константных (CH1, CH2, СНЗ) (см. рис. 1.30). Каждый домен иммуноглобулина имеет β-складчатую суперструктуру, в которой два остатка цистеина соединены дисульфидной связью.
Между двумя константными доменами СН1 и СН2 имеется участок, содержащий большое число остатков пролина, которые препятствуют формированию вторичной структуры и взаимодействию соседних Н-цепей на этом отрезке. Эта шарнирная область придает молекуле антитела гибкость. Между вариабельными доменами тяжелых и легких цепей находятся два идентичных антигенсвязывающих участка (активные центры для связывания антигенов), поэтому такие антитела часто называют бивалентами. В связывании антигена с антителом участвует не вся аминокислотная последовательность вариабельных участков обеих цепей, а всего лишь 20-30 аминокислот, расположенных в гипервариабельных областях каждой цепи. Именно эти области определяют уникальную способность каждого вида антитела взаимодействовать с соответствующим комплементарным антигеном.
Антитела - одна из линий защиты организма против внедрившихся чужеродных организмов. Их функционирование можно разделить на два этапа: первый этап - узнавание и связывание антигена на поверхности чужеродных организмов, что возможно благодаря наличию в структуре антитела антигенсвязывающих участков; второй этап - инициация процесса инактивации и разрушения антигена. Специфичность второго этапа зависит от класса антител. Существует пять классов тяжелых цепей, отличающихся друг от друга по строению константных доменов: α, δ, ε, γ и μ, в соответствии с которыми различают пять классов иммуноглобулинов: A, D, Е, G и М.
Особенности строения тяжелых цепей придают шарнирным участкам и С-концевым областям тяжелых цепей характерную для каждого класса конформацию. После связывания антигена с антителом конформационные изменения константных доменов определяют путь удаления антигена.
 Рис. 1. 30. Доменное строение IgG
Рис. 1. 30. Доменное строение IgG
Иммуноглобулины М
Иммуноглобулины М имеют две формы.
Мономерная форма - 1-й класс антител, продуцируемый развивающимся В-лимфоцитом. Впоследствии многие В-клетки переключаются на выработку других классов антител, но с тем же антигенсвязывающим участком. IgM встраивается в мембрану и выполняет роль антигенраспознающего рецептора. Встраивание IgM в мембрану клеток возможно благодаря наличию в хвостовой части участка 25 гидрофобных аминокислотных остатков.
Секреторная форма IgM содержит пять мономерных субъединиц, связанных друг с другом дисульфидными связями и дополнительной полипептидной J-цепью (рис. 1.31). Тяжелые цепи мономеров этой формы не содержат гидрофобной хвостовой части. Пентамер имеет 10 центров связывания с антигеном и поэтому эффективен в распознавании и удалении впервые попавшего в организм антигена. Секреторная форма IgM - основной класс антител, секретируемых в кровь при первичном иммунном ответе. Связывание IgM с антигеном изменяет конформацию IgM и индуцирует связывание его с первым белковым компонентом системы комплемента (система комплемента - набор белков, участвующих в уничтожении антигена) и активацию этой системы. Если антиген расположен на поверхности микроорганизма, система комплемента вызывает нарушение целостности клеточной мембраны и гибель бактериальной клетки.
Иммуноглобулины G
В количественном отношении этот класс иммуноглобулинов преобладает в крови (75% от всех Ig). IgG - мономеры, основной класс антител, секретируемый в кровь при вторичном иммунном ответе. После взаимодействия IgG с поверхностными антигенами микроорганизмов комплекс антиген-антитело способен связывать и активировать белки системы комплемента или может взаимодействовать со специфическими рецепторами макрофагов и нейтрофилов. Взаимодействие с фагоцитами приводит
 Рис. 1.31. Строение секреторной формы IgM
Рис. 1.31. Строение секреторной формы IgM
к поглощению комплексов антиген-антитело и разрушению их в фагосомах клеток. IgG - единственный класс антител, которые способны проникать через плацентарный барьер и обеспечивать внутриутробную защиту плода от инфекций.
Иммуноглобулины А
Основной класс антител, присутствующий в секретах (молоке, слюне, секретах дыхательных путей и кишечного тракта). IgA секретируются в основном в димерной форме, где мономеры связаны друг с другом через дополнительную J-цепь (рис. 1.32).
IgA не взаимодействуют с системой комплемента и фагоцитирующими клетками, но, связываясь с микроорганизмами, антитела препятствуют их присоединению к эпителиальным клеткам и проникновению в организм.
Иммуноглобулины Е
Иммуноглобулины Е представлены мономерами, в которых тяжелые ε-цепи содержат, так же как и μ-цепи иммуноглобулинов М, один вариабельный и четыре константных домена. IgE после секреции связываются своими
 Рис. 1.32. Строение IgA
Рис. 1.32. Строение IgA
С-концевыми участками с соответствующими рецепторами на поверхности тучных клеток и базофилов. В результате они становятся рецепторами для антигенов на поверхности данных клеток (рис. 1.33).
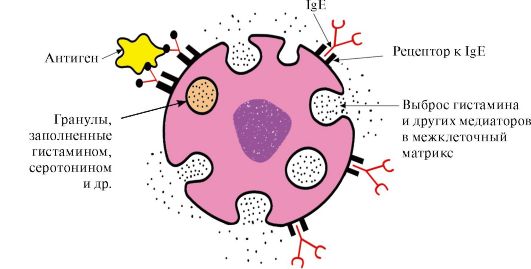 Рис. 1.33. Взаимодействие IgE с антигеном на поверхности тучной клетки
Рис. 1.33. Взаимодействие IgE с антигеном на поверхности тучной клетки
После того как происходит присоединение антигена к соответствующим антигенсвязывающим участкам IgE, клетки получают сигнал к секреции биологически активных веществ (гистамина, серотонина), которые в большой мере ответственны за развитие воспалительной реакции и за проявление таких аллергических реакций, как астма, крапивница, сенная лихорадка.
Иммуноглобулины D
Иммуноглобулины D обнаружены в сыворотке в очень небольшом количестве, они являются мономерами. В тяжелых δ-цепях имеются один вариабельный и три константных домена. IgD выполняют роль рецепторов В-лимфоцитов, другие функции пока неизвестны. Взаимодействие специфических антигенов с рецепторами на поверхности В-лимфоцитов (IgD) приводит к передаче этих сигналов в клетку и включению механизмов, обеспечивающих размножение данного клона лимфоцитов.
ТЕМА 1.7. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА БЕЛКОВ И МЕТОДЫ ИХ РАЗДЕЛЕНИЯ
1. Индивидуальные белки различаются по физико-химическим свойствам:
Форме молекул;
Молекулярной массе;
Суммарному заряду, величина которого зависит от соотношения анионных и катионных групп аминокислот;
Соотношению полярных и неполярных радикалов аминокислот на поверхности молекул;
Степени устойчивости к воздействию различных денатурирующих агентов.
2. Растворимость белков зависит от свойств белков, перечисленных выше, а также от состава среды, в которой растворяется белок (значения рН, солевого состава, температуры, наличия других органических веществ, способных взаимодействовать с белком). Величина заряда белковых молекул - один из факторов, влияющих на их растворимость. При потере заряда в изоэлектрической точке белки легче агрегируют и выпадают в осадок. Это особенно характерно для денатурированных белков, у которых на поверхности оказываются гидрофобные радикалы аминокислот.
На поверхности белковой молекулы имеются как положительно, так и отрицательно заряженные радикалы аминокислот. Количество этих групп, а следовательно, и суммарный заряд белков зависят от рН среды, т.е. соотношения концентрации Н+- и ОН - -групп. В кислой среде повышение концентрации Н+ приводит к подавлению диссоциации карбоксильных групп -СОО - + Н+ > - СООН и понижению отрицательного заряда белков. В щелочной среде связывание избытка ОН - протонами, образующимися при диссоциации аминогрупп -NH 3 + + ОН - - NH 2 + Н 2 О с образованием воды, приводит к уменьшению положительного заряда белков. Значение рН, при котором белок имеет суммарный нулевой заряд, называется изоэлектрической точкой (ИЭТ). В ИЭТ число положительно и отрицательно заряженных групп одинаково, т.е. белок находится в изоэлектрическом состоянии.
3. Разделение индивидуальных белков. Особенности строения и функционирования организма зависят от набора белков, синтезирующихся в нем. Изучение строения и свойств белков невозможно без их выделения из клетки и очистки от других белков и органических молекул. Стадии выделения и очистки индивидуальных белков:
Разрушение клеток изучаемой ткани и получение гомогената.
Разделение гомогената на фракции центрифугированием, получение ядерной, митохондриальной, цитозольной или иной фракции, содержащей искомый белок.
Избирательная тепловая денатурация - кратковременное нагревание раствора белков, при котором можно удалить часть денатурированных белковых примесей (в том случае, если белок относительно термостабилен).
Высаливание. Различные белки выпадают в осадок при разных концентрациях соли в растворе. Постепенно повышая концентрацию соли, можно получить ряд отдельных фракций с преимущественным содержанием выделяемого белка в одной из них. Наиболее часто для фракционирования белков используют сульфат аммония. Белки с наименьшей растворимостью выпадают в осадок при небольших концентрациях солей.
Гель-фильтрация - метод просеивания молекул через набухшие гранулы сефадекса (трехмерные полисахаридные цепи декстрана, имеющие поры). Скорость прохождения белков через колонку, заполненную сефадексом, будет зависеть от их молекулярной массы: чем меньше масса молекул белка, тем легче они проникают внутрь гранул и дольше там задерживаются, чем больше масса, тем быстрее они элюируют с колонки.
Ультрацентрифугирование - метод, заключающийся в том, что белки в центрифужной пробирке помещают в ротор ультрацентрифуги. При вращении ротора скорость оседания белков пропорциональна их молекулярной массе: фракции более тяжелых белков расположены ближе ко дну пробирки, более легкие - ближе к поверхности.
Электрофорез - метод, в основе которого лежат различия в скорости движения белков в электрическом поле. Эта величина пропорциональна заряду белков. Электрофорез белков проводят на бумаге (в этом случае скорость движения белков пропорциональна только их заряду) или в полиакриламидном геле с определенной величиной пор (скорость движения белков пропорциональна их заряду и молекулярной массе).
Ионообменная хроматография - метод фракционирования, основанный на связывании ионизированных групп белков с противоположно заряженными группами ионообменных смол (нерастворимых полимерных материалов). Прочность связывания белка со смолой пропорциональна заряду белка. Белки, адсорбированные на ионообменном полимере, можно смыть растворами NaCl с возрастающими концентрациями; чем меньше заряд белка, тем меньшая концентрация NaCl потребуется, чтобы смыть белок, связанный с ионогенными группами смолы.
Аффинная хроматография - наиболее специфический метод выделения индивидуальных белков.К инертному полимеру ковалентно присоединяется лиганд какого-либо белка. При пропускании раствора белков через колонку с полимером за счет комплементарного связывания белка с лигандом на колонке адсорбируется только специфичный для данного лиганда белок.
Диализ - метод, применяемый для удаления низкомолекулярных соединений из раствора выделяемого белка. Метод основан на неспособности белков проходить через полупроницаемую мембрану в отличие от низкомолекулярных веществ. Применяется для очистки белков от низкомолекулярных примесей, например от солей после высаливания.
ЗАДАНИЯ ДЛЯ ВНЕАУДИТОРНОЙ РАБОТЫ
1. Заполните табл. 1.4.
Таблица 1.4. Сравнительный анализ структуры и функций родственных белков - миоглобина и гемоглобина
а) вспомните строение активного центра Мв и НЬ. Какую роль играют гидрофобные радикалы аминокислот в формировании активных центров этих белков? Опишите строение активного центра Мв и НЬ и механизмы присоединения к нему лигандов. Какую роль играют остатки Гис F 8 и Гис Е 7 в функционировании активного центра Мв иНв?
б) какими новыми свойствами по сравнению с мономерным миоглобином обладает близко родственный олигомерный белок - гемоглобин? Объясните роль кооперативных изменений конформации протомеров в молекуле гемоглобина, влияние концентраций СО 2 и протонов на сродство гемоглобина к кислороду, а также роль 2,3-БФГ в аллостерической регуляции функции НЬ.
2. Дайте характеристику молекулярным шаперонам, обращая внимание на связь их структуры с функцией.
3. Какие белки объединяют в семейства? На примере семейства иммуноглобулинов определите сходные черты строения и родственные функции белков этого семейства.
4. Часто для биохимических и медицинских целей требуются очищенные индивидуальные белки. Объясните, на каких физико-химических свойствах белков основаны используемые методы их разделения и очистки.
ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ
1. Выберите правильные ответы.
Функции гемоглобина:
A. Транспорт О 2 из легких в ткани Б. Транспорт Н + из тканей в легкие
B. Поддержание постоянства рН крови Г. Транспорт СО 2 из легких в ткани
Д. Транспорт СО 2 из тканей в легкие
2. Выберите правильные ответы. Лигандом α-протомера Нв является: A. Гем
Б. Кислород
B. СО Г. 2,3-БФГ
Д. β-Протомер
3. Выберите правильные ответы.
Гемоглобин в отличие от миоглобина:
A. Имеет четвертичную структуру
Б. Вторичная структура представлена только α-спиралями
B. Относится к сложным белкам
Г. Взаимодействует с аллостерическим лигандом Д. Ковалентно связан с гемом
4. Выберите правильные ответы.
Сродство Нв к О 2 уменьшается:
A. При присоединении одной молекулы О 2 Б. При отщеплении одной молекулы О 2
B. При взаимодействии с 2,3-БФГ
Г. При присоединении к протомерам Н + Д. При снижении концентрации 2,3-БФГ
5. Установите соответствие.
Для типов Нв характерно:
A. В дезокси форме образует фибриллярные агрегаты Б. Содержит в составе две α- и две δ-цепи
B. Преобладающая форма Нв в эритроцитах взрослого человека Г. В активном центре содержит гем с Fе+ 3
Д. Содержит в составе две α- и две γ-цепи 1. НвА 2.
6. Установите соответствие.
Лиганды Нв:
A. Связывается с Нв в аллостерическом центре
Б. Имеет очень высокое сродство к активному центру Нв
B. Присоединяясь, повышает сродство Нв к O 2 Г. Окисляет Fe+ 2 в Fе+ 3
Д. Образует ковалентную связь с гисF8
7. Выберите правильные ответы.
Шапероны:
A. Белки, присутствующие во всех отделах клетки
Б. Синтез усиливается при стрессовых воздействиях
B. Участвуют в гидролизе денатурированных белков
Г. Участвуют в поддержании нативной конформации белков
Д. Создают органеллы, в которых формируется конформация белков
8. Установите соответствие. Иммуноглобулины:
A. Секреторная форма имеет пентамерную форму
Б. Класс Ig, проникающих через плацентарный барьер
B. Ig - рецептор тучных клеток
Г. Основной класс Ig, присутствующих в секретах эпителиальных клеток. Д. Рецептор В-лимфоцитов, активация которого обеспечивает размножение клеток
9. Выберите правильные ответы.
Иммуноглобулины Е:
A. Вырабатываются макрофагами Б. Имеют тяжелые ε-цепи.
B. Встраиваются в мембрану Т-лимфоцитов
Г. Выполняют роль мембранных рецепторов антигенов на тучных клетках и базофилах
Д. Ответственны за проявление аллергических реакций
10. Выберите правильные ответы.
Метод разделения белков основан на различиях в их молекулярной массе:
A. Гель-фильтрация
Б. Ультрацентрифугирование
B. Электрофорез в полиакриламидном геле Г. Ионообменная хроматография
Д. Аффинная хроматография
11. Выберите правильный ответ.
Метод разделения белков основан на различиях в их растворимости в воде:
A. Гель-фильтрация Б. Высаливание
B. Ионообменная хроматография Г. Аффинная хроматография
Д. Электрофорез в полиакриламидном геле
ЭТАЛОНЫ ОТВЕТОВ К «ЗАДАНИЯМ ДЛЯ САМОКОНТРОЛЯ»
1. А, Б, В, Д
2. А, Б, В, Д
5. 1-В, 2-А, 3-Г
6. 1-В, 2-Б, 3-А
7. А, Б, Г, Д
8. 1-Г; 2-Б, 3-В
ОСНОВНЫЕ ТЕРМИНЫ И ПОНЯТИЯ
1. Олигомерные белки, протомер, четвертичная структура белков
2. Кооперативные изменения конформации протомеров
3. Эффект Бора
4. Аллостерическая регуляция функций белков, аллостерический центр и аллостерический эффектор
5. Молекулярные шапероны, белки теплового шока
6. Семейства белков (сериновые протеазы, иммуноглобулины)
7. IgM-, G-, Е-, А-связь структуры с функцией
8. Суммарный заряд белков, изоэлектрическая точка белков
9. Электрофорез
10. Высаливание
11. Гель-фильтрация
12. Ионообменная хроматография
13. Ультрацентрифугирование
14. Аффинная хроматография
15. Электрофорез белков плазмы крови
ЗАДАНИЯ ДЛЯ АУДИТОРНОЙ РАБОТЫ
1. Сравните зависимости степеней насыщения гемоглобина (Нв) и миоглобина (Mb) кислородом от его парциального давления в тканях
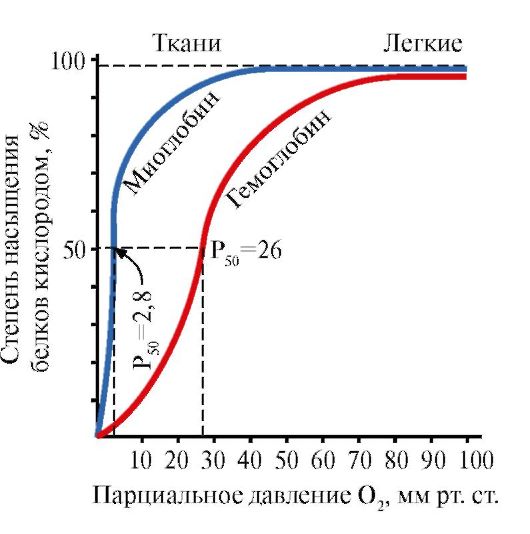 Рис. 1.34. Зависимость насыщения Мв и
НЬ
кислородом от его парциального давления
Рис. 1.34. Зависимость насыщения Мв и
НЬ
кислородом от его парциального давления
Обратите внимание, что форма кривых насыщения белков кислородом различна: для миоглобина - гипербола, для гемоглобина - сигмоидная форма.
1. сравните значения парциального давления кислорода, при котором Мв и Нв насыщены О 2 на 50%. Какой из этих белков характеризуется более высоким сродством к О 2 ?
2. какие особенности строения Мв определяют его высокое сродство к О 2 ?
3. какие особенности строения Нв позволяют ему отдавать О 2 в капиллярах покоящихся тканей (при относительно высоком парциальном давлении О 2) и резко увеличивать эту отдачу в работающих мышцах? Какое свойство олигомерных белков обеспечивает этот эффект?
4. вычислите, какое количество О 2 (в %) отдает оксигенированный гемоглобин покоящейся и работающей мышце?
5. сделайте выводы о связи структуры белка с его функцией.
2. Количество освобождаемого гемоглобином кислорода в капиллярах зависит от интенсивности процессов катаболизма в тканях (эффект Бора). Как изменения метаболизма в тканях регулируют сродство НЬ к O 2 ? Влияние СО 2 и Н+ на сродство НЬ к О 2
 1. опишите эффект Бора.
1. опишите эффект Бора.
2. в каком направлении протекает процесс, представленный на схеме:
а) в капиллярах легких;
б) в капиллярах тканей?
3. в чем физиологическое значение эффекта Бора?
4. почему взаимодействие Нв с Н+ на участках, удаленных от гема, изменяет сродство белка к О 2 ?
3. Сродство НЬв к О 2 зависит от концентрации его лиганда - 2,3-бифосфо- глицерата, который является аллостерическим регулятором сродства Нв к О 2 . Почему взаимодействие лиганда в участке, удаленном от активного центра, влияет на функцию белка? Как 2,3-БФГ регулирует сродство НЬ к О 2 ? Для решения задачи ответьте на следующие вопросы:
1. где и из чего синтезируется 2.3-бифосфоглицерат (2,3-БФГ)? Напишите его формулу, укажите заряд данной молекулы.
2. с какой формой гемоглобина (окси или дезокси) взаимодействует БФГ и почему? В каком участке молекулы Нв происходит взаимодействие?
3. в каком направлении протекает процесс, представленный на схеме
а) в капиллярах тканей;
б) в капиллярах легких?
4. где должна быть более высокой концентрация комплекса
Нв-2,3-БФГ:
а) в капиллярах мышц, находящихся в состоянии покоя,
б) в капиллярах работающих мышц (при условии одинаковой концентрации БФГ в эритроцитах)?
5. как изменится сродство Нв к кислороду при адаптации человека к условиям высокогорья, если концентрация БФГ в эритроцитах при этом повышается? В чем физиологическое значение этого явления?
4. Разрушение 2,3-БФГ при хранении консервированной крови нарушает функции Нв. Как изменится сродство Нв к O 2 в консервированной крови, если концентрация 2,3-БФГ в эритроцитах может уменьшатся с 8 до 0,5 ммол/л. Можно ли переливать такую кровь тяжелобольным пациентам, если концентрация 2,3-БФГ восстанавливается не ранее чем через трое суток? Можно ли добавлением в кровь 2,3-БФГ восстановить функции эритроцитов?
5. Вспомните строение простейших молекул иммуноглобулинов. Какую роль играют иммуноглобулины в работе иммунной системы? Почему Ig часто называют бивалентами? Как структура Ig связана с их функцией? (Опишите на примере какого-либо класса иммуноглобулинов.)
Физико-химические свойства белков и методы их разделения.
6. Как суммарный заряд белка влияет на его растворимость?
а) определите суммарный заряд пептида при рН 7
Ала-Глу-Тре-Про-Асп-Лиз-Цис
б) как изменится заряд этого пептида при рН >7, рН <7, рН <<7?
в) что такое изоэлектрическая точка белка (ИЭТ) и в какой среде лежит
ИЭТ данного пептида?
г) при каком значении рН будет наблюдаться наименьшая растворимость данного пептида.
7. Почему кислое молоко в отличие от свежего при кипячении «сворачивается» (т.е. белок молока казеин выпадает в осадок)? В свежем молоке молекулы казеина имеют отрицательный заряд.
8. Для разделения индивидуальных белков используется метод гельфильтрации. Смесь, содержащую белки А, В, С с молекулярными массами, равными соответственно 160 000, 80 000 и 60 000, анализировали методом гель-фильтрации (рис. 1.35). Гранулы набухшего геля проницаемы для белков с молекулярной массой меньше 70 000. Какой принцип лежит в основе данного метода разделения? Какой из графиков правильно отражает результаты фракционирования? Укажите порядок выхода белков А, В и С с колонки.
 Рис. 1.35. Использование метода гель-фильтрации для разделения белков
Рис. 1.35. Использование метода гель-фильтрации для разделения белков
9. На рис. 1.36, А представлена схема электрофореза на бумаге белков сыворотки крови здорового человека. Относительные количества белковых фракций, полученных с помощью этого метода, составляют: альбумины 54-58%, α 1 -глобулины 6-7%, α 2 -глобулины 8-9%, β-глобулины 13%, γ-глобулины 11-12%.
 Рис. 1.36 Электрофорез на бумаге белков плазмы крови здорового человека (А) и больного (Б)
Рис. 1.36 Электрофорез на бумаге белков плазмы крови здорового человека (А) и больного (Б)
I - γ-глобулины; II - β-глобулины; III - α 2 -глобулин; IV - α 2 -глобулин; V - альбумины
Многие болезни сопровождаются количественными изменениями в составе сывороточных белков (диспротеинемии). Характер этих изменений учитывается при постановке диагноза и оценке тяжести и стадии заболевания.
С помощью данных, приведенных в табл. 1.5, сделайте предположение о заболевании, для которого характерен электрофоретический профиль, представленный на рис. 1.36.
Таблица 1.5. Изменение концентрации белков сыворотки крови при патологии
Доказано существование 4 уровней структурной организации белковой молекулы.
Первичная структура белка – последовательность расположения аминокислотных остатков в полипептидной цепи. В белках отдельные аминокислоты связаны друг с другом пептидными связями , возникающими при взаимодействии a-карбоксильных и a-аминогрупп аминокислот.
К настоящему времени расшифрована первичная структура десятков тысяч разных белков. Для определения первичной структуры белка методами гидролиза выясняют аминокислотный состав. Затем определяют химическую природу концевых аминокислот. Следующий этап - определение последовательности аминокислот в полипептидной цепи. Для этого используют избирательный частичный (химический и ферментативный) гидролиз. Возможно применение рентгеноструктурного анализа, а также данных о комплементарной нуклеотидной последовательности ДНК.
Вторичная структура белка – конфигурация полипептидной цепи, т.е. способ упаковки полипептидной цепи в определенную конформацию. Процесс этот протекает не хаотично, а в соответствии с программой, заложенной в первичной структуре.
Стабильность вторичной структуры обеспечивается в основном водородными связями, однако определенный вклад вносят ковалентные связи – пептидные и дисульфидные.
Наиболее вероятным типом строения глобулярных белков считают a-спираль . Закручивание полипептидной цепи происходит по часовой стрелке. Для каждого белка характерна определенная степень спирализации. Если цепи гемоглобина спирализованы на 75%, то пепсина-всего на 30%.
Тип конфигурации полипептидных цепей, обнаруженных в белках волос, шелка, мышц, получил название b-структуры . Сегменты пептидной цепи располагаются в один слой, образуя фигуру, подобную листу, сложенному в гармошку. Слой может быть образован двумя или большим количеством пептидных цепей.
В природе существуют белки, строение которых не соответствует ни β-, ни a-структуре, например, коллаген - фибриллярный белок, составляющий основную массу соединительной ткани в организме человека и животных.
Третичная структура белка – пространственная ориентация полипептидной спирали или способ укладки полипептидной цепи в определенном объеме. Первый белок, третичная структура которого была выяснена рентгеноструктурным анализом - миоглобин кашалота (рис. 2).
В стабилизации пространственной структуры белков, помимо ковалентных связей, основную роль играют нековалентные связи (водородные, электростатические взаимодействия заряженных групп, межмолекулярные ван-дер-ваальсовы силы, гидрофобные взаимодействия и т.д.).
По современным представлениям, третичная структура белка после завершения его синтеза формируется самопроизвольно. Основной движущей силой является взаимодействие радикалов аминокислот с молекулами воды. При этом неполярные гидрофобные радикалы аминокислот погружаются внутрь белковой молекулы, а полярные радикалы ориентируются в сторону воды. Процесс формирование нативной пространственной структуры полипептидной цепи называют фолдингом . Из клеток выделены белки, названные шаперонами. Они участвуют в фолдинге. Описан ряд наследственных заболеваний человека, развитие которых связывают с нарушением вследствие мутаций процесса фолдинга (пигментозы, фиброзы и др.).
Методами рентгеноструктурного анализа доказано существование уровней структурной организации белковой молекулы, промежуточных между вторичной и третичной структурами. Домен - это компактная глобулярная структурная единица внутри полипептидной цепи (рис. 3). Открыто много белков (например, иммуноглобулины), состоящих из разных по структуре и функциям доменов, кодируемых разными генами.
Все биологические свойства белков связаны с сохранностью их третичной структуры, которую называют нативной . Белковая глобула не является абсолютно жесткой структурой: возможны обратимые перемещения частей пептидной цепи. Эти изменения не нарушают общей конформации молекулы. На конформацию молекулы белка оказывают влияние рН среды, ионная сила раствора, взаимодействие с другими веществами. Любые воздействия, приводящие к нарушению нативной конформации молекулы, сопровождаются частичной или полной потерей белком его биологических свойств.
Четвертичная структура белка - способ укладки в пространстве отдельных полипептидных цепей, обладающих одинаковой или разной первичной, вторичной или третичной структурой, и формирование единого в структурном и функциональном отношениях макромолекулярного образования.
Белковую молекулу, состоящую из нескольких полипептидных цепей, называют олигомером , а каждую входящую в него цепь - протомером . Олигомерные белки чаще построены из четного числа протомеров, например, молекула гемоглобина состоит из двух a- и двух b-полипептидных цепей (рис. 4).
Четвертичной структурой обладает около 5% белков, в том числе гемоглобин, иммуноглобулины. Субъединичное строение свойственно многим ферментам.
Белковые молекулы, входящие в состав белка с четвертичной структурой, образуются на рибосомах по отдельности и лишь после окончания синтеза образуют общую надмолекулярную структуру. Биологическую активность белок приобретает только при объединении входящих в его состав протомеров. В стабилизации четвертичной структуры принимают участие те же типы взаимодействий, что и в стабилизации третичной.
Некоторые исследователи признают существование пятого уровня структурной организации белков. Это метаболоны - полифункциональные макромолекулярные комплексы разных ферментов, катализирующих весь путь превращений субстрата (синтетазы высших жирных кислот, пируватдегидрогеназный комплекс, дыхательная цепь).
МИНИСТЕРСТВО КУЛЬТУРЫ, ОБРАЗОВАНИЯ И ЗДРАВООХРАНЕНИЯ
РЕСПУБЛИКИ КАЗАХСТАН
ПАВЛОДАРСКИЙ УНИВЕРСИТЕТ
КАФЕДРА БИОЛОГИИ
КОНТРОЛЬНАЯ РАБОТА
Предмет: «Биохимия»
Выполнила ст-ка
г. Павлодар, 2004г.
1. Вода в живых организмах. Строение и свойства воды.
2. Структурные формулы пуриновых и пиримидиновых оснований, входящих в состав нуклеиновых кислот.
3. Свойства ферментов, специфичность действий ферментов. Отличия денатурированного белка от нативного.
4. Витамин D, витамеры этого витамина. Признаки авитаминоза D. Природные источники витамина D.
5. Схема дихотомического распада D-глюкозы (гликолиз).
6. Структурная формула пептида-валил-изолейцил-метионил-аргенин.
Все живое на нашей планете на 2/3 состоит из воды. На первом месте в живом веществе по массе стоят микроорганизмы, на втором-растения, на третьем-животные, на последнем - человек. Бактерии на 81 проц. состоят из воды, споры-на 50 проц., ткани животных в среднем на 70 проц., лимфа - 90 проц., в крови содержится около 79 проц. Самая богатая водой ткань - стекловидное тело глаза, которое содержит до 99 проц. влаги, самая бедная - зубная эмаль - всего лишь 0,2 проц.
Вода в организме выполняет несколько функций: растворенные в ней вещества реагируют друг с другом, вода помогает удалению отходов обмена веществ, служит регулятором температуры, являясь хорошим переносчиком тепла, а также смазочным веществом.
У живых организмов вода может синтезироваться в тканях. Так, например, у верблюда жир в горбу, окисляясь, может дать до 40 л воды. Человек, выпивая 2,5 л воды в сутки, ежедневно промывает желудок 10 л жидкостей и испаряет 0,7 л воды.
Изучение химического состава клеток показывает, что в живых организмах нет никаких особых химических элементов, свойственных только им: именно в этом проявляется единство химического состава живой и неживой природы.
Велика роль химических элементов в клетке: N и S входят в состав белков, Р - в ДНК и РНК, Mg - в состав многих ферментов и молекулу хлорофилла, Сu - компонент многих окислительных ферментов, Zn- гормона поджелудочной железы, Fe - молекулы гемоглобина, I - гормона тироксина и т. д. Наиболее важны для клетки анионы НРО42-, Н2РО4-, СО32-, Сl-, НСОз- и катионы Na+, К+, Ca2+
Содержание катионов и анионов в клетке отличается от их концентрации в среде, окружающей клетку, вследствие активной регуляции переноса веществ мембраной. Так обеспечивается постоянство химического состава живой клетки. С гибелью клетки концентрация веществ в среде и в цитоплазме выравнивается. Из неорганических соединений важное значение имеют вода, минеральные соли, кислоты, основания.
Вода в функционирующей клетке занимает до 80% ее объема и находится в ней в
двух формах: свободной и связанной. Молекулы связанной воды прочно соединены с
белками и образуют вокруг них водные оболочки, изолирующие белки друг от друга.
Полярность молекул воды, способность образовывать водородные связи объясняет ее
высокую удельную теплоемкость. Вследствие этого в живых системах предотвращаются
резкие колебания температуры, происходит распределение и отдача тепла в клетке.
Благодаря связанной воде клетка способна выдерживать низкие температуры. Ее
содержание в клетке составляет примерно 5%, и 95% приходится на свободную воду.
Последняя растворяет многие вещества, вовлекаемые клеткой в обмен.
В высокоактивных клетках, например в ткани головного
мозга, на долю воды приходится около 85%, а в мышцах-более 70%; в менее
активных клетках, например в жировой ткани, вода составляет около 40% ее массы.
В. живых организмах вода не только растворяет многие вещества; с ее участием
происходят реакции гидролиза - расщепления органических соединений до промежуточных
и конечных веществ.
Вещество
Поступление в клетку
Местонахождение и преобразование
Свойства
У растений - из окружающей среды; у животных образуется
непосредственно в клетке при
углеводов и поступает из окружающей среды
В цитоплазме, вакуолях, матриксе органелл, ядерном соке, клеточной стенке, межклетниках. Вступает в реакции синтеза, гидролиза и окисления
Растворитель. Источник кислорода, осмотический регулятор, среда для
физиологических и биохимических процессов,
химический компонент, терморегулор
Стоит отметить, что различные органические вещества при своем окислении образуют различные количества воды. Чем богаче молекула органического вещества водородом, тем больше образуется при его окислении воды. При окислении 100 г жира образуется 107 мл воды, 100 г углеводов - 55 мл воды, 100 г белков - 41 мл воды.
Суточная потребность организма человека в воде составляет около 40 г воды на 1 кг веса. У детей грудного возраста потребность в воде на 1 кг веса в три - четыре раза выше, чем у взрослых.
Вода в организмах живых существ не только выполняет транспортную функцию, она также используется в процессах обмена веществ. Включение воды в органические вещества в большом масштабе имеет место у зеленых растений, у которых при использовании солнечной энергии из воды, углекислого газа и минеральных азотистых веществ синтезируются углеводы, белки, липиды и иные органические вещества.
Поступление воды в организм регулируется чувством жажды. Уже при первых признаках сгущения крови в результате рефлекторного возбуждения определенных участков коры головного мозга возникает жажда - стремление к питью. При потреблении даже большого количества воды единовременно, кровь не обогащается водой сразу, не разжижается. Объясняется это тем, что вода из крови быстро поступает в межклеточные пространства и увеличивает количество межклеточной воды. Всосавшаяся в кровь и отчасти в лимфу из кишечника вода в значительной части поступает в кожу и на некоторое время там задерживается. В печени также удерживается некоторое количество поступившей в организм воды.
Вода выделяется из организма, главным образом, почками, с мочой, в небольшом количестве ее выделяют стенки кишечника, затем потовые железы (через кожу) и легкие с выдыхаемым воздухом. Количество воды, выделяемой из организма не постоянно. При сильном потении из организма с потом может выделяться 5 и более литров воды в сутки. В этом случае количество воды, выделяемой почками, уменьшается, моча сгущается. Уменьшается выделение мочи при ограничении питья. Однако сгущение мочи возможно до определенного предела, и при дальнейшем ограничении питья задерживается выведение из организма конечных продуктов азотистого обмена и минеральных веществ, что отрицательно отражается на жизнедеятельности организма. При обильном поступлении воды в организм, выделение мочи увеличивается.
Вода в природе. Вода - весьма распространенное на Земле вещество. Почти 3 4 поверхности земного шара покрыты водой, бразующей океаны, моря, реки и озера. Много воды находится в газообразном состоянии в виде паров в атмосфере; в виде огромных масс снега и льда лежит она круглый год на вершинах высоких гор и в полярных странах. В недрах земли также находится вода, пропитывающая почву и горные породы.
Вода имеет очень большое значение в жизни растений, живот ых и человека. Согласно современным представлениям, само происхождени жизни связывае ся с морем. Во всяком организме вода представля т собой среду, в которой протекают химические процессы, обеспечивающие жизнедеятельность организма; кроме ого, она сама принимает участие в целом ряде биохимических реакций.
Чистая вода представляет собой есцветную прозрачную жидкость. Плотность воды при переходе ее из твердого состояния в жидкое не уменьшается, как почти у всех других веществ, а возрастает. При нагревании воды от 0 до 4 С плотность ее также увеличивается. При 4 С вода имеет максимальную плотность, и лишь при дальнейшем нагревании ее плотность уменьшается.
Большое значение в жизни природы имеет и тот факт, что вода. обладает аномально высокой теплоемкостью , Поэтому в ночное время, а также при переходе от лета к зиме вода остывает медленно, а днем или при переходе от зимы к лету так же медленно нагревается, являясь, таким образом, регулятором температуры на земном шаре.
Молекула воды имеет угловое строение; входящие в ее состав ядра образуют равнобедренный треугольник, в основании которого находятся два протона, а в вершине - ядро атома кислорода, Межъядерные расстояния О- близки к 0,1 нм, расстояние между ядрами атомов водорода равно примерно 0,15 нм. И восьм электронов, составляющих внешний электронный слой атома кислорода в молекуле воды
Вода - весьма реакционноспособное вещество. Оксиды многих металлов и неметаллов соединяются с водой, образуя основания и кислоты; некоторые соли образуют с водой кристаллогидраты; наиболее активные металлы взаимодействуют с водой с выделением водорода.
Вода обладает также каталитической способностью. В отсутствие следов влаги практически не протекают некоторые обычные реакции; например, хлор не взаимодействует с металлами, фтороводород не разъедает стекло, натрий не окисляется в атмосферы воздуха.
Вода способна соединяться с рядом веществ, находящихся при обычных условиях в газообразном состоянии, образуя при этом так: называемые гидраты газов. Примерами могут служить соединения Хе 6Н О, CI 8H O, С Н 6Н О, С Н 17Н О, которые выпадают в виде кристаллов при температурах от 0 до 24 °С (обычно при повышенном давлении соответствующего газа). Подобные соединения возникают в результате заполнения молекулами газа (“гостя”) межмолекулярных полостей, имеющихся в структуре воды (“хозяина”); они называются соединениями включения или клатратами.
Пуриновые нуклеозиды:
Пиримидиновые нуклеозиды:
ФЕРМЕНТЫ, органические вещества белковой природы, которые синтезируются в клетках и во много раз ускоряют протекающие в них реакции, не подвергаясь при этом химическим превращениям. Вещества, оказывающие подобное действие, существуют и в неживой природе и называются катализаторами. Ферменты (от лат. fermentum – брожение, закваска) иногда называют энзимами (от греч. en – внутри, zyme – закваска). Все живые клетки содержат очень большой набор ферментов, от каталитической активности которых зависит функционирование клеток. Практически каждая из множества разнообразных реакций, протекающих в клетке, требует участия специфического фермента. Изучением химических свойств ферментов и катализируемых ими реакций занимается особая, очень важная область биохимии – энзимология.
Многие ферменты находятся в клетке в свободном состоянии, будучи просто растворены в цитоплазме; другие связаны со сложными высокоорганизованными структурами. Есть и ферменты, в норме находящиеся вне клетки; так, ферменты, катализирующие расщепление крахмала и белков, секретируются поджелудочной железой в кишечник. Секретируют ферменты и многие микроорганизмы.
Первые данные о ферментах были получены при изучении процессов брожения и пищеварения. Большой вклад в исследование брожения внес Л.Пастер, однако он полагал, что соответствующие реакции могут осуществлять только живые клетки. В начале 20 в. Э.Бухнер показал, что сбраживание сахарозы с образованием диоксида углерода и этилового спирта может катализироваться бесклеточным дрожжевым экстрактом. Это важное открытие послужило стимулом к выделению и изучению клеточных ферментов. В 1926 Дж.Самнер из Корнеллского университета (США) выделил уреазу; это был первый фермент, полученный в практически чистом виде. С тех пор обнаружено и выделено более 700 ферментов, но в живых организмах их существует гораздо больше. Идентификация, выделение и изучение свойств отдельных ферментов занимают центральное место в современной энзимологии.
Ферменты, участвующие в фундаментальных процессах превращения энергии, таких, как расщепление сахаров, образование и гидролиз высокоэнергетического соединения аденозинтрифосфата (АТФ), присутствуют в клетках всех типов – животных, растительных, бактериальных. Однако есть ферменты, которые образуются только в тканях определенных организмов. Так, ферменты, участвующие в синтезе целлюлозы, обнаруживаются в растительных, но не в животных клетках. Таким образом, важно различать «универсальные» ферменты и ферменты, специфичные для тех или иных типов клеток. Вообще говоря, чем более клетка специализирована, тем больше вероятность, что она будет синтезировать набор ферментов, необходимый для выполнения конкретной клеточной функции.
Ферменты как белки. Все ферменты являются белками, простыми или сложными (т.е. содержащими наряду с белковым компонентом небелковую часть). См. также БЕЛКИ.
Ферменты – крупные молекулы, их молекулярные массы лежат в диапазоне от 10 000 до более 1 000 000 дальтон (Да). Для сравнения укажем мол. массы известных веществ: глюкоза – 180, диоксид углерода – 44, аминокислоты – от 75 до 204 Да. Ферменты, катализирующие одинаковые химические реакции, но выделенные из клеток разных типов, различаются по свойствам и составу, однако обычно обладают определенным сходством структуры.
Структурные особенности ферментов, необходимые для их функционирования, легко утрачиваются. Так, при нагревании происходит перестройка белковой цепи, сопровождающаяся потерей каталитической активности. Важны также щелочные или кислотные свойства раствора. Большинство ферментов лучше всего «работают» в растворах, pH которых близок к 7, когда концентрация ионов H+ и OH- примерно одинакова. Связано это с тем, что структура белковых молекул, а следовательно, и активность ферментов сильно зависят от концентрации ионов водорода в среде.
Не все белки, присутствующие в живых организмах, являются ферментами. Так, иную функцию выполняют структурные белки, многие специфические белки крови, белковые гормоны и т.д.
Коферменты и субстраты. Многие ферменты с большой молекулярной массой проявляют каталитическую активность только в присутствии специфических низкомолекулярных веществ, называемых коферментами (или кофакторами). Роль коферментов играют большинство витаминов и многие минеральные вещества; именно поэтому они должны поступать в организм с пищей. Витамины РР (никотиновая кислота, или ниацин) и рибофлавин, например, входят в состав коферментов, необходимых для функционирования дегидрогеназ. Цинк – кофермент карбоангидразы, фермента, катализирующего высвобождение из крови диоксида углерода, который удаляется из организма вместе с выдыхаемым воздухом. Железо и медь служат компонентами дыхательного фермента цитохромоксидазы.
Вещество,
подвергающееся превращению в присутствии фермента, называют субстратом.
Субстрат присоединяется к ферменту, который ускоряет разрыв одних химических
связей в его молекуле и создание других; образующийся в результате продукт
отсоединяется от фермента. Этот процесс представляют следующим образом:
Механизм действия ферментов. Скорость ферментативной реакции зависит от концентрации субстрата [S] и количества присутствующего фермента. Эти величины определяют, сколько молекул фермента соединится с субстратом, и именно от содержания фермент-субстратного комплекса зависит скорость реакции, катализируемой данным ферментом. В большинстве ситуаций, представляющих интерес для биохимиков, концентрация фермента очень мала, а субстрат присутствует в избытке. Кроме того, биохимики исследуют процессы, достигшие стационарного состояния, при котором образование фермент-субстратного комплекса уравновешивается его превращением в продукт.
Выяснение механизмов действия ферментов во всех деталях – дело будущего, однако некоторые важные их особенности уже установлены. Каждый фермент имеет один или несколько активных центров, с которыми и связывается субстрат. Эти центры высокоспецифичны, т.е. «узнают» только «свой» субстрат или близкородственные соединения. Активный центр формируют особые химические группы в молекуле фермента, ориентированные друг относительно друга определенным образом. Происходящая так легко потеря ферментативной активности связана именно с изменением взаимной ориентации этих групп. Молекула субстрата, связанного с ферментом, претерпевает изменения, в результате которых разрываются одни и образуются другие химические связи. Чтобы этот процесс произошел, необходима энергия; роль фермента состоит в снижении энергетического барьера, который нужно преодолеть субстрату для превращения в продукт. Как именно обеспечивается такое снижение – до конца не установлено.
Ферментативные реакции и энергия. Высвобождение энергии при метаболизме питательных веществ, например при окислении шестиуглеродного сахара глюкозы с образованием диоксида углерода и воды, происходит в результате последовательных согласованных ферментативных реакций. В животных клетках в превращениях глюкозы в пировиноградную кислоту (пируват) или молочную кислоту (лактат) участвуют 10 разных ферментов. Этот процесс называется гликолизом. Первая реакция – фосфорилирование глюкозы – требует участия АТФ. На превращение каждой молекулы глюкозы в две молекулы пировиноградной кислоты расходуются две молекулы АТФ, но при этом на промежуточных этапах из аденозиндифосфата (АДФ) образуются 4 молекулы АТФ, так что весь процесс в целом дает 2 молекулы АТФ.
Далее пировиноградная кислота окисляется до диоксида углерода и воды при участии ферментов, ассоциированных с митохондриями. Эти превращения образуют цикл, называемый циклом трикарбоновых кислот, или циклом лимонной кислоты. См. также МЕТАБОЛИЗМ.
Окисление одного вещества всегда сопряжено с восстановлением другого: первое отдает атом водорода, а второе его присоединяет. Катализируют эти процессы дегидрогеназы, обеспечивающие перенос атомов водорода от субстратов к коферментам. В цикле трикарбоновых кислот одни специфические дегидрогеназы окисляют субстраты с образованием восстановленной формы кофермента (никотинамиддинуклеотида, обозначаемого НАД), а другие окисляют восстановленный кофермент (НАДЧН), восстанавливая другие дыхательные ферменты, в том числе цитохромы (железосодержащие гемопротеины), в которых атом железа попеременно то окисляется, то восстанавливается. В конечном итоге восстановленная форма цитохромоксидазы, одного из ключевых железосодержащих ферментов, окисляется кислородом, попадающим в наш организм с вдыхаемым воздухом. Когда происходит горение сахара (окисление кислородом воздуха), входящие в его состав атомы углерода непосредственно взаимодействуют с кислородом, образуя диоксид углерода. В отличие от горения, при окислении сахара в организме кислород окисляет собственно железо цитохромоксидазы, но в конечном итоге его окислительный потенциал используется для полного окисления сахаров в ходе многоступенчатого процесса, опосредуемого ферментами.
На отдельных этапах окисления энергия, заключенная в питательных веществах, высвобождается в основном маленькими порциями и может запасаться в фосфатных связях АТФ. В этом принимают участие замечательные ферменты, которые сопрягают окислительные реакции (дающие энергию) с реакциями образования АТФ (запасающими энергию). Этот процесс сопряжения известен как окислительное фосфорилирование. Не будь сопряженных ферментативных реакций, жизнь в известных нам формах была бы невозможна.
Ферменты выполняют и множество других функций. Они катализируют разнообразные реакции синтеза, включая образование тканевых белков, жиров и углеводов. Для синтеза всего огромного множества химических соединений, обнаруженных в сложных организмах, используются целые ферментные системы. Для этого нужна энергия, и во всех случаях ее источником служат фосфорилированные соединения, такие, как АТФ.
Ферменты и пищеварение. Ферменты – необходимые участники процесса пищеварения. Только низкомолекулярные соединения могут проходить через стенку кишечника и попадать в кровоток, поэтому компоненты пищи должны быть предварительно расщеплены до небольших молекул. Это происходит в ходе ферментативного гидролиза (расщепления) белков до аминокислот, крахмала до сахаров, жиров до жирных кислот и глицерина. Гидролиз белков катализирует фермент пепсин, содержащийся в желудке. Ряд высокоэффективных пищеварительных ферментов секретирует в кишечник поджелудочная железа. Это трипсин и химотрипсин, гидролизующие белки; липаза, расщепляющая жиры; амилаза, катализирующая расщепление крахмала. Пепсин, трипсин и химотрипсин секретируются в неактивной форме, в виде т.н. зимогенов (проферментов), и переходят в активное состояние только в желудке и кишечнике. Это объясняет, почему указанные ферменты не разрушают клетки поджелудочной железы и желудка. Стенки желудка и кишечника защищает от пищеварительных ферментов и слой слизи. Некоторые важные пищеварительные ферменты секретируются клетками тонкого кишечника.
Большая часть энергии, запасенной в растительной пище, такой, как трава или сено, сосредоточена в целлюлозе, которую расщепляет фермент целлюлаза. В организме травоядных животных этот фермент не синтезируется, и жвачные, например крупный рогатый скот и овцы, могут питаться содержащей целлюлозу пищей только потому, что целлюлазу вырабатывают микроорганизмы, заселяющие первый отдел желудка – рубец. С помощью микроорганизмов происходит переваривание пищи и у термитов.
Ферменты находят применение в пищевой, фармацевтической, химической и текстильной промышленности. В качестве примера можно привести растительный фермент, получаемый из папайи и используемый для размягчения мяса. Ферменты добавляют также в стиральные порошки.
Ферменты в медицине и сельском хозяйстве. Осознание ключевой роли ферментов во всех клеточных процессах привело к широкому их применению в медицине и сельском хозяйстве. Нормальное функционирование любого растительного и животного организма зависит от эффективной работы ферментов. В основе действия многих токсичных веществ (ядов) лежит их способность ингибировать ферменты; таким же эффектом обладает и ряд лекарственных препаратов. Нередко действие лекарственного препарата или токсичного вещества можно проследить по его избирательному влиянию на работу определенного фермента в организме в целом или в той или иной ткани. Например, мощные фосфорорганические инсектициды и нервно-паралитические газы, разработанные в военных целях, оказывают свой губительный эффект, блокируя работу ферментов – в первую очередь холинэстеразы, играющей важную роль в передаче нервного импульса.
Чтобы лучше понять механизм действия лекарственных препаратов на ферментные системы, полезно рассмотреть, как работают некоторые ингибиторы ферментов. Многие ингибиторы связываются с активным центром фермента – тем самым, с которым взаимодействует субстрат. У таких ингибиторов наиболее важные структурные особенности близки к структурным особенностям субстрата, и если в реакционной среде присутствуют и субстрат и ингибитор, между ними наблюдается конкуренция за связывание с ферментом; при этом чем больше концентрация субстрата, тем успешнее он конкурирует с ингибитором. Ингибиторы другого типа индуцируют в молекуле фермента конформационные изменения, в которые вовлекаются важные в функциональном отношении химические группы. Изучение механизма действия ингибиторов помогает химикам создавать новые лекарственные препараты.
Гликолиз.
Гликолиз является первым, а в анаэробных условиях основным этапом на пути «пользования глюкозы и других углеводов для обеспечения биоэнергетических потребностей живых организмов. Кроме того, на промежуточных стадиях гликолиза образуются трехуглеродные фрагменты, используемые- для биосинтеза ряда веществ.
Стержневым этапом гликолиза
является окислительная деструкция глюкозы до двух молекул пирувата - соли пировиноградной
кислоты с использованием и качестве
окислителя двух молекул NAD. Стереометрическое уравнение процесса записывается в виде:
1. Превращение глюкозы в
глюкозо-6-фосфат, катализируемое гексокиназои:
2. Изомеризация
глюкозо-6-фосфата во фруктозо-6-фосфат, катализируемая глюкозо-6-фосфат изомеразой:
3. Фосфорилированне фруктозо-б-фосфата
до фруктозо-1,6-дифосфата, катализируемое
6-фосфофруктокнназой:
4. Распад
фруктозо-1,6-дпфосфата на глпцсральдегпд-3-фосфат и дигмдрокси-ацетонфосфат, катализируемый фру ктозод и
фосфат альдолазой:
5. Изомеризация дигидроксиацетонфосфата в глицеральдегид-З-фосфат,
катализируемая триозофосфат
изомеразой:
Если последующие стадии являются преобладающим путем.превращения глюкозы, то эта реакция обеспечивает постепенное прекращение дигидрокс-ацетонфосфата в глицеральдегид-3-фосфат.
6. Окисление глицеральдегид-3-фосфата до 1,3-дифосфаглицерата, катализируемое глицералъдегид-З-фосфат дегидрсиеназой:
Процесс происходит через промежуточное образование триэфира
между окисляемой альдегидной группой и Sll-группой
остатка Цпстеппа, «ходящего к активный центр фермента. Эта
связь затем подвергается фосфоролнзу неорганическим фосфатом с регенерацией активного центра и образованием смешанного ангидрида 3-фосфоглицершювой п фосфорной кислот:
7. Перенос фосфата с
1,3-дпфосфогл|щерата па АДФ с образованием молекулы АТФ, катализируемый фосфоглицерат
киназой (название дано в соответствии с обратной реакцией):
8. Изомеризация
3-фосфоглпцерата в 2-фосфоглпцерат, катализируемая фос-фоглицерат
мутазой:
9. Дегидратация
2-фосфоглпцерата, катализируемая еиолазой п приводящая к образованию
сильного макроэрга - фосфоеиолппрувата:
10. Перенос фосфата от
фосфоенол пиру вата па АДФ с образованием еще одной молекулы АТФ, катализируемый
пируоат киназой (название дано в соответствии с обратной реакцией):
Прежде чем просуммировать эти уравнения, следует обратить
внимание на то] обстоятельство, что на первых
стадиях гликолиза расходуется две макроэргические связи в молекулах А"ГФ для превращения глюкозы в глюкозо-6-фосфат и фруктозо-6-фосфата во фруктозо-1,6-дифосфат. На
последующих стадиях в расчете на одну
исходную молекулу глюкозы две молекулы АДФ фосфорилируются в реакции и две в реакции. Таким образом, итогом является превращение двух молекул
АДФ и двух молекул ортофосфата в две молекулы АТФ. С учетом этого
суммарное уравнение следует записать в виде:
Если вести отсчет от
глюкозо-6-фосфата, то уравнение примет вид:
Схема гликолиза
(превращение глюкозы в две молекулы пирувата)
Нативный и денатурированный белок.
Белки и нуклеиновые кислоты в живых организмах образуются путем последовательного наращивания полимерной цепи мономерными звеньями, порядок присоединения которых определяется программирующими биосинтез нуклеиновыми кислотами. Однако последние сами по себе определяют лишь первичную структуру создаваемого биополимера. Для того чтобы биополимер принял необходимую для его функционирования пространственную структуру нативную структуру, необходимо, чтобы последняя была запрограммирована самой первичной структурой белка.
Нативность белка обуславливает тритичная структура. Нативный белок – это белок, способный выполнять все биологические функции. Тритичная структура легко разрушается, за счет изменения pH среды, изменения температуры, солей тяжелых металлов и т.д. Белок теряет свои свойства по мере повышения температуры, неизбежно наступает момент, когда нативная структура становится термодинамически неустойчивой. Ее разрушение приводит к тому, что полипептидная цепь теряет упорядоченную конфирмацию и превращается в полимер с непрерывно изменяющейся пространственной структурой. В химии высокомолекулярных соединений такие образования называют статистическим клубком. В биохимии превращение нативного белка в статистический клубок называют денатурацией белка.
Денатурированный белок лишен всякой биологической активности и в биологических системах может я основном быть использован лишь как источник аминокислот, т.е. как продукт питания.
Обратное превращение
денатурированного белка в нативной возможно лишь в случае, когда нативная структура запрограммирована
в первичной структуре.
Витамины
группы
D
.
Известно около десяти витаминов D, незначительно различающихся между собой по строению. Все они относятся к группе стероидов - сложных органических соединений с конденсированными кольцами. Все витамины группы D участвуют в управлении процессом отложения кальция и фосфора в растущих костях человека. При отсутствии витаминов D этот процесс нарушается, в результате чего кости становятся мягкими и деформируются. Такое явление называется рахитом и свойственно только детскому возрасту.
Витамины D содержатся в некоторых продуктах питания, но в количестве, недостаточном для роста человека. Недостающее количество витаминов D организм восполняет за счет имеющегося в организме 7-дегидро-холестерина - соединения из группы стероидов, близкого по строению к витаминам D. Содержащийся непосредственно под кожей человека 7-дегидрохолестерин под действием солнечных лучей превращается в витамин D3:
Витамин D (кальциферол] очень
близок по строению к
витамину D3 и образуется из стероидного спирта- эргостерина,
содержащегося в дрожжах, плесени и др. также под действием облучения.
Структурная формула
пептида-валил-изолейцил-метионил-аргенин.
Библиография
1. Д.Э., Техника и производство. М., 1972г
2. Хомченко Г.П. , Химия для поступающих в ВУЗы. М., 1995г.
3. Прокофьев М.А., Энциклопедический словарь юного химика. М., 1982г.
4. Глинка Н.Л., Общая химия. Ленинград, 1984г.
5. Ахметов Н.С., Неорганическая химия. Москва, 1992г.